INTRODUCTION
The Korean bullhead (Pseudobagrus fulvidraco; Fig. 1) is a type of fish with over 205 species within the family of Bagridae, the order of Siluiformes that lives mainly in the fresh water area of the continents of Asia and the parts of Africa (Kang & Lee, 1996). The Korean bullhead lives under the water of mud of a river of gradual slope. The Korean bullhead is a nocturnal fish that hides mainly under rocks and starts being active at night, and it lives off of small animals such as the eggs of other fish, prawn. The egg laying season for the Korean bullhead is from mid May to mid July and this species takes 2 to 3 years for the matured eggs to spawning (Lim et al., 2008).

Within Korea, the Korean bullhead lives mainly in the rivers that flow to the Yellow Sea. It was usually not seen in the rivers that flow to the East Sea or the South Sea, however, recently, the habitat of the Korean bullhead is spreading as the Korean bullhead has been sited at the areas near Nakdong River and Milyang River, Gyeongsangnam-do, Korea (Kang & Lee, 1996). Also, the Korean bullhead has been known to be spectacular in taste that it is widely used as the main ingredient of the fresh-water spicy fish stew such as catfishes or carps, and its demand is also increasing. Recently, the development of culture method of bullhead resulted in the huge seed production and culture in the Jeollabuk-do and Chungcheongnam-do, Korea (Shin et al., 2000).
The inhibition of the first cleavage is a method of chromosome engineering. Chromosome manipulation can accomplish a short term maximization in the increase of productivity of culture (Mair, 1993). The inhibition of The first cleavage is used in the induction of the tetraploid of the chromosome manipulation, the mitotic gynogenetic diploid, and the androgenetic diploid; at this time, for the efficient inhibition of the first cleavage, the time of inhibition of the first cleavage that depends on the water temperature must be measured (Mair, 1993).
The induction of the parthenogenesis of vertebrates has been induced by poking the unfertilized eggs with a needle (Park et al., 2006). Afterwards, dealing with the cold shock has been decided to be effective at inducing the parthenogenesis by suppressing the first mitosis, which is the first cleavage in amphibians (Park et al., 2006). Also, the doubling of the genetic materials of gynogenetic haploid is possible by suppressing the second meiosis and the first cleavage; The suppression of the second meiosis is possible by dealing with the temperature, water pressure, and chemicals that are induced from triploid for certain fish, and the suppression of the first cleavage is used by a combination of the treatment of high pressure and water pressure when inducing tetraploid (Komen et al., 1991). Streisinger et al. (1981) induced homozygous mitotic gynogenetic diploid zebrafish (Zebra danios) via dealing with high temperature and hydrostatic pressure at the first mitosis, and Onozato (1984) produced mitotic gynogenetic diploid in salmon via dealing with hydrostatic pressure. When inducing gynogenetic diploid, the effective suppression of the second polar body release and the first cleavage can be different depending on the type of fish used, pressure applied, and time exposed (Thorgaard & Allen, 1987). Since the Dettlaff unit (τ0) denotes the period of one mitotic interval during the cleavage that occurs simultaneously in the beginning or the separation between two continuous cell divisions (Shelton et al., 1997), the τ0 differs depending on the temperature, through the regression, the relationship between the τ0 and temperature is predictable developmental stages within similar reproductive species (Dettlaff, 1986).
Until now, the τ0 has been used to estimate the best time to manipulate chromosomes for the following species: paddlefish (Polyodon spathula), shovelnose sturgeon (Scaphirhynchus platorynchus), carp (Cyprinus carpio), tench (Tinca tinca), black crappie (Pomoxis nigromaculatus), far eastern catfish (Silurus asotus), winter flounder (Pseudopleuronectes americanus), greenling (Hexagrammos otakii), black plaice (Pleuronectes obscurus), and Korean rose bitterling (Rhodeus uyekii) (Flajšhans et al., 1993; Shelton & Rothbard, 1993; Mims et al., 1997; Shelton et al., 1997; Gomelsky et al., 2000; Park & Im, 2001; Park & Johnson, 2002; Park et al., 2006; Park & Im, 2010; Kim et al., 2011).
To this day, not much regarding research related to chromosome manipulation of Korean bullhead is known; only results by Kang & Lee (1996), Park et al. (2003), and Park et al. (2004) on early life history and sex hormone of Korean bullhead, sex differentiation, sex exchange, growth effects by tamoxifen are known. Thus, this paper conducted research relating the time of the first cleavage and the τ0 that can be used as basic data for effective manipulation of chromosomes of Korean bullhead.
MATERIALS AND METHODS
The broodstock of Korean bullhead (Pseudobagrus fulvidraco) used in this experiment were collected from the areas of the Kum River, Kangkyung-eup, Nonsan city, Chungcheongnam-do, Korea from April to June 2012 and were transferred to National Fisheries Research and Development Institute, Inland Aquaculture Research Center to be used. The average length and weight of the fish parental generation were measured with a digital vernier caliper (CD-20CP, Japan) and a digital balance (JW-1, Korea) to the tenths in grams. With references to Baek et al. (2000) and Hwang et al. (1999), 10 IU/g BW of human chorionic gonadotropin (HCG; Sigma, St. Louis, MO, USA) was injected to the broodstock fish. After a 24 hour, collection of the egg and sperm was done by pressing the abdominals and the eggs were hatched by artificial fertilization through the dry method.
In order to examine the reproductive characteristics of the parental generation, gonadosomatic index (GSI) and hepatosomatic index (HSI) of 20 female and 20 male fish were measured. GSI and HSI were calculated by the following formulas: GSI (%)=(weight of reproductive organs/weight of fish)×100, HSI (%)=(weight of liver/weight of fish)×100. Moreover, the number of eggs per unit of weight and diameters of fertilized eggs of the females were measured. The number of eggs per unit of weight was adjusted to the number of eggs per 10 g of weight and the number of eggs was calculated by dividing the weight of a fertilized egg by the average weight of egg. The diameters of fertilized eggs were measured by the eyepiece micrometer of a microscope (Axioskop, Zeiss, Germany) under 50× magnification. Fifty fertilized eggs were measured and averaged.
To investigate the time of the first cleavage of fertilized eggs depending on the difference between temperatures of egg development, also known as the Dettlaff unit (mitotic interval, τ0) of different cleavage frequencies and water temperatures, eggs were fertilizes at 21, 24, 27, and 30 and were sampled °C every 5 min after fertilization. Each sample was fixed in 10% neutral formalin (100 ml formalin, 6.5 g Na2HPO4 ∙ 12H2O, 4 g KH2PO4, fresh water 900 ml) and kept at 4°C until observation. τ0 was examined under microscope under 50X magnification. For τ0, the time it took for about 10% of the entire eggs to reach 2-cell stage (τI) and 8-cell stage (τIII) at each hatching temperature were measured according to the method of Ignatyeva (1975). Average τ0 was calculated by the formula τ0=(τIII - τI)/2. Linear regression analysis was performed to find the relationship between the average τ0 and water temperature. This experiment was performed in triplicate.
RESULTS AND DISCUSSION
The average length and weight of broodstock of Korean bullhead (Pseudobagus fulvidraco) were 11.9±1.81 cm and 64.1±2.87 g (n=20) for females and 9.4±1.16 cm and 49.9±1.06 g (n=20) for males (Table 1). Gonadosomatic index (GSI) for females was 10 times greater than that for males as it was 20.2±1.43% for females and 2.3± 0.78% for males, and hepatosomatic index (HSI) for females was 20% smaller than that for males as it was 1.0±0.49% for females and 0.8±0.19% for males (Table 1). The average number of eggs was 1,573±425 eggs/10 g egg weight.
| Body length (cm) | Body weight (g) | GSI (%) | HSI (%) | |
|---|---|---|---|---|
| Female (n=20) | 11.9±1.81 | 64.1±2.87 | 20.2±1.43 | 1.0±0.49 |
| Male (n=20) | 09.4±1.16 | 49.9±1.06 | 2.3±0.78 | 0.8±0.19 |
In Baek et al. (2000) when they inserted luteinizing hormone-releasing hormone analogue (LHRHa) pellet into females of spotted halibut (Verasper variegates), they found no signs of final maturation of fertilized eggs or ovulation inducement, but when they injected human chorionic gonadotropin (HCG) of 300 IU/kg BW, the most effective results of final maturation and ovulation inducement were observed. Moreover, it has been reported that it is possible to mass collect the eggs with high hatching rate when 1,000 IU/kg BW of HCG was injected to females of rabbitfish (Siganus canaliculatus) (Hwang et al., 1999). In this study, when preliminary experiments were done based on results from Baek et al. (2000) and Hwang et al. (1999), the final maturation and the amount of ovulation induced in the ovaries of females of Korean bullhead were higher when treated with HCG than with LHRHa pellet. This is consistent with results from Baek et al. (2000). Also, the optimal HCG turned out to be 10,000 IU/kg BW and this is higher than what is found in Hwang et al. (1999). Therefore, in order to improve the accuracy of the study, HCG was used rather than LHRHa that is often used in spawning inducement in the parental females.
The fertilized eggs of Korean bullhead are spherically separative with transparent membrane. The egg has mucous layer on the surface so that attachment can be easily done. Also, the egg yolks are light yellow without oil globules (Kang & Lee, 1996). The radius of Korean bullhead egg was similar to what is found in Kang & Lee, (1996) (1.5±0.06 mm, n=30), but was smaller than that of far eastern catfish (Silutus asotus) or Korean rose bitterling (Rhodeus uyekii) (3.8±0.04 mm) and bigger than that of black plaice (Pleuronectes obscurus) (0.8± 0.01 mm) (Park & Im, 2010; Kim et al., 2011).
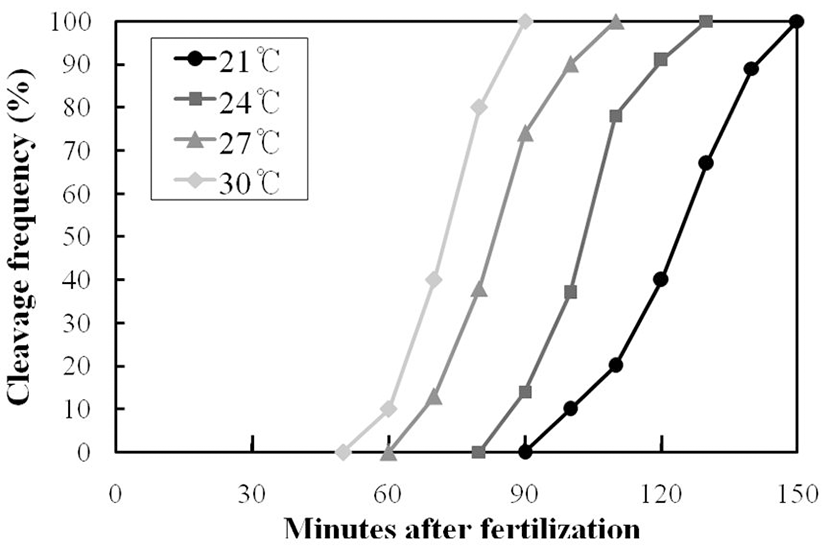
Figure 2 shows the results the first cleavage time distributions according to the time passes after fertilization of Korean bullhead egg with the different water temperatures. At 21°C water temperatures of the egg development, the first cleavage started at 90 min after fertilization, the frequency of cleavage increased gradually and appeared the 50% frequency of first cleavage at 120~125 min after fertilization, and was completed at 150 min after fertilization. Therefore, it appeared that it took 60 min for the first cleavage to be completed. At 24°C water temperature of the egg development, the first cleavage started at 80 min after fertilization, and the frequency of cleavage increased gradually and was completed at 125 min after fertilization. It took 45 min for the first cleavage to be completed and 100~105 min after fertilization for the 50% frequency of the first cleavage to show up. During the egg development, at 27°C water temperature the first cleavage started at 60 min after fertilization and finished at 105 min after fertilization, which means it took 45 min for it to be completed. The 50% frequency of the first cleavage appeared at 80~85 min after fertilization. At 30°C water temperature of egg development, the first cleavage started at 50 min after fertilization and was completed at 80 min after fertilization. It took 30 min for the first cleavage to be completed and 75~80 min for the 50% frequency of the first cleavage to show up.
In the development of Korean bullhead egg, the mitotic interval was 33.4±2.08 min at 21°C water temperature, 31.5±3.06 min at 24°C water temperature, 28.1±2.11 min at 27 water temperature, °C and 26.4± 3.35 min at 30°C water temperature (Fig. 3). In the developing Korean bullhead egg, as shown in Fig. 3, mitotic interval time (τ0; Y) according to the increase of water temperature decreased linearly and showed a linear equation Y=–1.13X+58.15 (R2=0.98, n=30) according to water temperature (X).
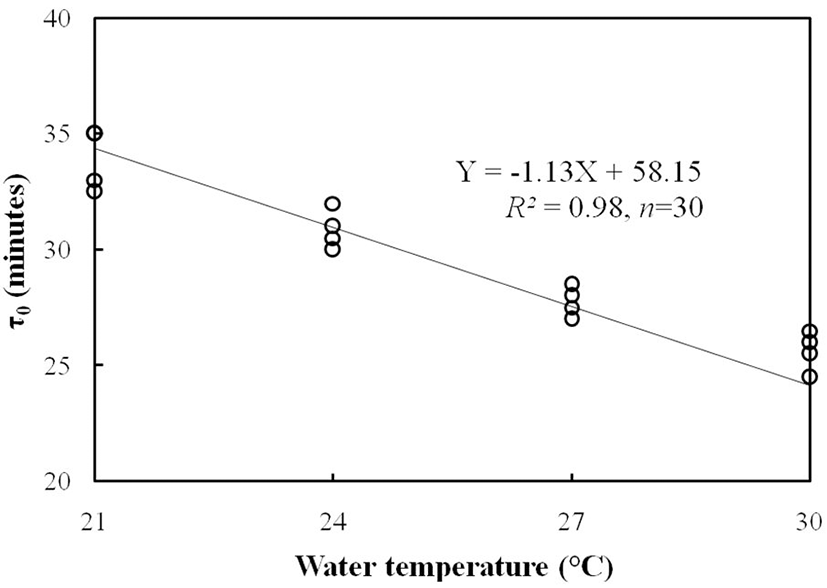
In this experiment, the times to 1 cell stage, 4 cell stage and 16 cell stage were excluded since there were some difficulties in deciding the accurate time of initiating one cell stage, in determining the starting time of the first cleavage and in measuring the time of synchronized cleavage after 8 cell stage. In egg development of Korean bullhead on the standard of fertilization time (for example, the initiation time of the first cleavage in fertilized eggs of Korean bullhead and the results obtained from the τ0), it showed the tendency that the initiation time of the first cleavage ran fast according to the water temperature and τ0 was reduced as well. Except for this study, there were similar results of the reduction of mitotic interval according to rising the water temperature in far eastern catfish, winter flounder (Pseudopleuronectes americanus), greenling (Hexagrammos otakii), black plaice, Korean rose bitterling (Park & Im, 2001; Park & Johnson, 2002; Park et al., 2006; Park & Im, 2010; Kim et al., 2011).
The initiation time of the first cleavage and the τ0 according to water temperature shown in this study determined the optimal treatment time for inhibition of the first cleavage and would be an effective foundation for chromosomal manipulation through production of the androgenetic diploid, the mitotic gynogenetic diploid and tetrapliod. According to Park et al. (2004), Korean bullhead go through sexual differentiation when they are about 2.5 cm in total body length and maturation of reproductive organs takes place faster in females than in males. Also, oral injection of tamoxifen and estradiol-17β at 3~9 days after hatching for 7 days resulted in virilization and feminization at 200 ppm and 40 ppm, respectively. Moreover, injection of tamoxifen at 50 mg/kg BW for 180 days shows that growth was faster and that more males than females were developed (sexual ratio of 7:3) than control group (Park et al., 2003, 2004). In conclusion, the results from this study, the early study of Park et al. (2003) and Park et al. (2004) would be useful in improving the accuracy of chromosomal manipulation through sex changes and in developing a way to enhance hatching and growth in larvae.