INTRODUCTION
Sevenband grouper, Epinephelus septemfasciatus is a fish genus in the Epinephelus subfamily of the family Serranidae, in the order of Perciformes. The subfamily, Epinephelinae contains about 159 species in 15 genera (FAO, 1993). Sevenband groupers, a carnivorous species, mostly inhabit near coral reefs in subtropical or tropical regions and are distributed widely along the southern coast of Korea and Jeju Island. It is also found along the southern coast of Japan, the South China Sea, and as far as the Indian Ocean (Kim et al., 2001). It is a commercially significant product for South Korea, Japan, China, and some Southeast Asian countries. It, along with the longtooth grouper, is considered a local Jeju culinary specialty. However, the sevenband grouper catch has been decreasing rapidly. In Korea the sevenband grouper spawns during summer season, and the embryogenesis and hatching rate are dependent on water temperate and salinity. However, little data are available regarding fertilized egg, embryonic development and larval stages. The techniques for seven-band grouper seedling production are needed for the Korean aquaculture industry to be more competitive and coastal resources management.
Some research on larval rearing, egg development, and the early life history of Epinephelinae were done in brown spotted grouper (E. tauvina) (Hussain & Higuchi, 1980), Nassau grouper (E. striatus) (Powell & Tucker, 1992), dusky grouper (E. marginatus) (Glamuzina et al., 1998), and longtooth grouper (E. bruneus) (Oh et al., 2003) for the aquaculture. Quality issues of cultured fish and advanced commercial production for cultural management were studied as well under various conditions such as: water temperature (Watanabe et al., 1995), rearing tank color, density (Duray et al., 1996), aeration, and light intensity (Toledo et al., 2002). This study aims to examine egg development, early larval development under controlled water temperature and salinity for seedling production and feeding management.
MATERIALS AND METHODS
After artificial fertilization, using eggs selected from mature females and sperm from mature males, we measured the egg size and oil globules and examined their growth and development in elapsed time progress. The experiments were carried out in a thermostatic chamber (22±0.5°C) and observed with a microscope (Zeiss, Germany) and a profile projector (Mitutoyco, Japan). Developmental progress, from the 8 cell stage, the 16 cell stage, morula stage, blastula stage, gastrula stage, Kupffer's vesicle occurrence, embryo formation, and hatching, was determined when each stage was 50% complete. The fertilized eggs were contained in 1.0 L beakers and experiments were repeated thrice.
To investigate the egg developmental progress depending on the water temperature, the experimental groups were maintained at 22°C and 25°C separately. Each fertilized group had 100 fertilized eggs and was contained in a 250 ml beaker. The time was measured for each developmental stage in the morula stage, blastula stage, gastrula stage, embryo formation, and hatching. The hatching rate for both hatched larvae and failed ones was calculated as day 1 from hatching. Also, results from artificial seawater at the salinity of 34 ppt, 26 ppt, and 18 ppt respectively, were examined to determine characteristics of egg development under controlled salinity. Each group had 50 fertilized eggs and was contained in a 100 ml beaker respectively, experiments were repeated thrice.
During the experiments, reared larval fish were fed rotifer Brachionus rotundiformis (size: 110 to 210 μm) in 15 individuals/ml. They were also fed approximately 500,000 cell/ml of Nannochloropsis oculata and Isochrysis galbana respectively. Larvae were fed an artificial diet (size: 150 μm. INVE, USA) since the fifteenth day after hatching and Artemia (2 to 3 per ml) with rotifer B. rotundiformis from the sixteenth day after hatching. The artificial diet’s quantity was increased according to the larval development (Fig. 1).
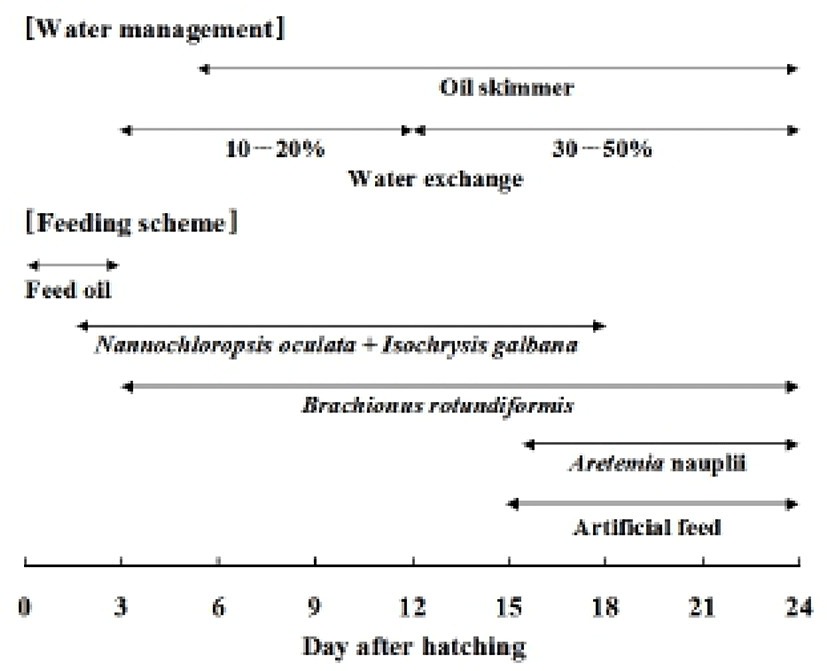
The yolk absorption rate and mouth opening time were examined at 22°C and 25°C. The yolk absorption rate and oil globule absorption rate of the larvae hatched from 2,000 to 3,000 reared eggs in a 20 L acrylic tank were investigated. Samples of 5 to 10 larvae were taken daily for measurements of yolk and oil globule. The specimens were anesthetized with MS-222 and then measured on a profile projector up to the nearest 1.0 μm (Fig. 2). The yolk and oil globule in each larvae were measured for volume with the Blaxter and Hempel method (1963) as follows:
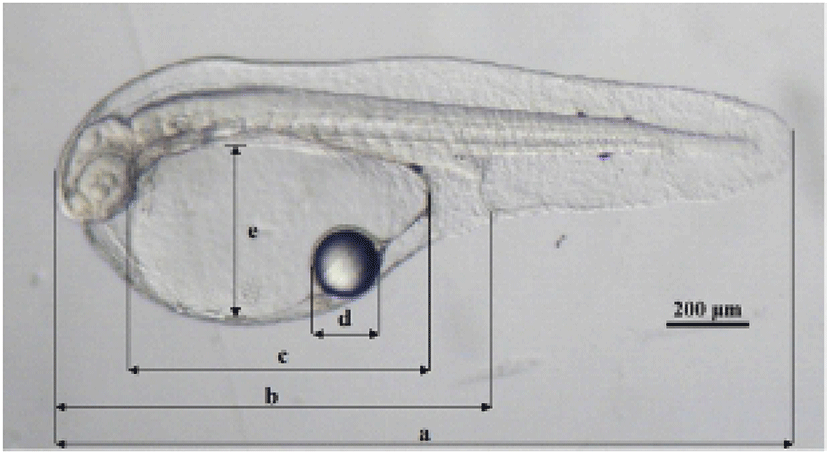
Yolk volume = π/6×Ih2
(I: major axis of yolk, h: minor axis of yolk)
Oil globule volume = π/6×d3
(d: diameter of oil globule)
The mouth opening was measured when 80% of mouth opened in the same method as the yolk absorption measurement.
The samples of hatched larvae, collected regularly from a 20 L acrylic tank and an indoor concrete water tank (7.0×7.0×1.0 m, effective water capacity of 35 ton) were analyzed to find size changes depending on its rearing condition and time progress. The samples were measured for diameter (d) with a Shirota formula (1970), with
The data was analyzed for statistical significance of means using the ANOVA-test and Duncan's multiple range test (Duncan, 1955) with SAS software.
RESULTS
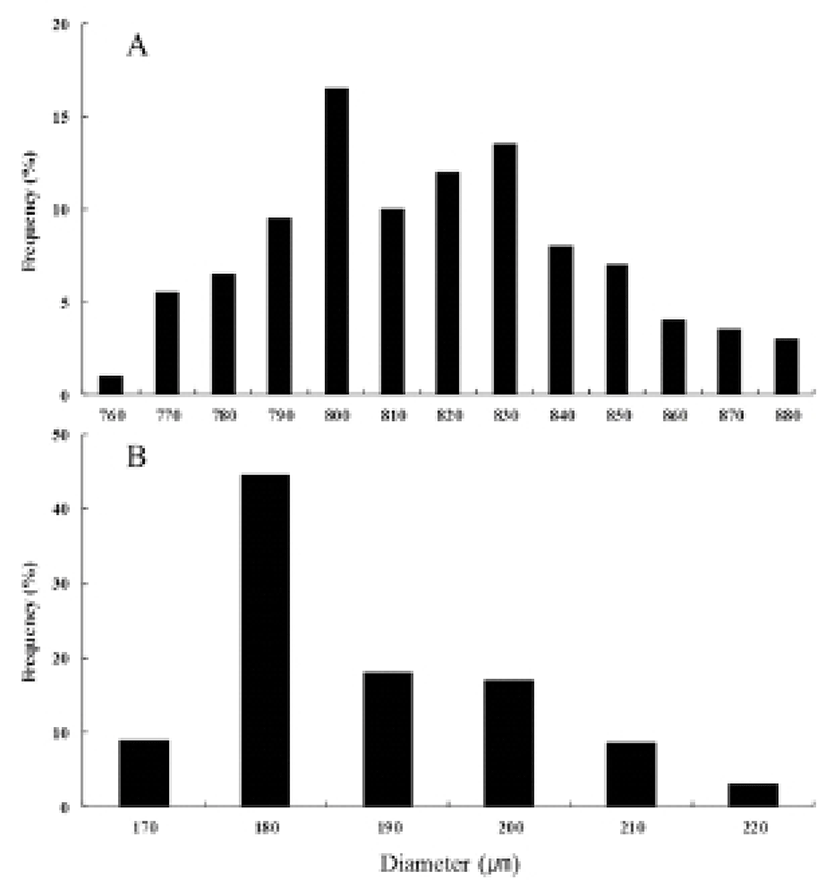
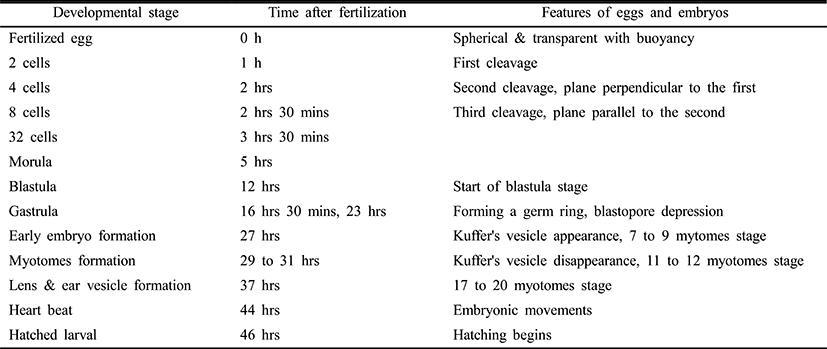
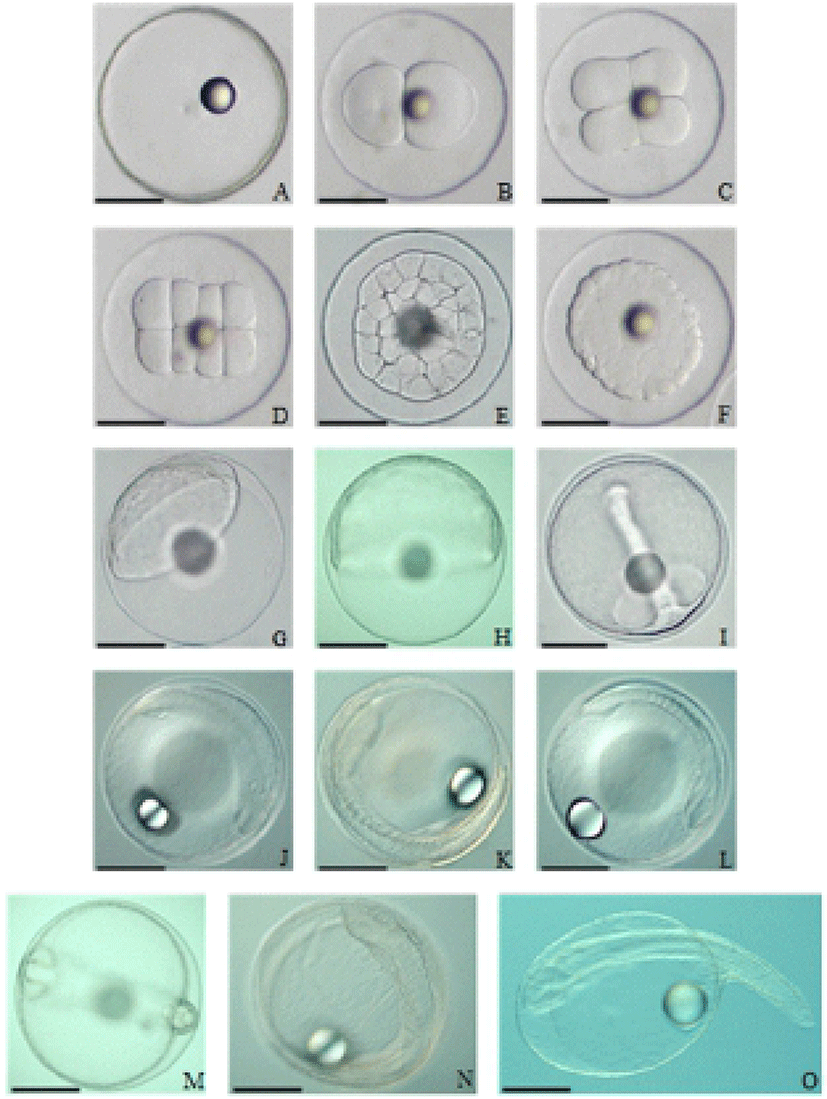
The fertilized eggs, with a diameter from 790 to 890 μm (an average diameter of 821.8±2.0 μm), were spherical and transparent with buoyancy (Fig. 3). Oil globules had a diameter range from 170 to 230 μm (an average diameter of 192.9±0.9 μm). Table 1 and Fig. 4 detail the phases of embryonic development at 22°C (Table 1, Fig. 4). Shortly after fertilization, perivitelline space and blastodisc were formed (Fig. 4A). One hour after fertilization, the blastodisc divided into 2 cells for the first cleavage (Fig. 4B). The eggs developed into 4 cells two hours after fertilization (Fig. 4C), 8 cells after 2 hours and 30 minutes (Fig. 4D), and 32 cells after 3 hours 30 minutes (Fig. 4E). Then blastomeres derived during the cleavages and formed the morula after 5 hours (Fig. 4F). The blastula stage started after 12 hours (Fig. 4G), forming the blastoderm. 16 hours after the fertilization, gastrulation started (Fig. 4H), forming a germ ring, as the blastoderm spread around the yolk. Then after 23 hours, embryonic cells began to invaginate, forming embryonic shield (Fig. 4I). 27 hours after the fertilization, the head divided forming optic vesicles, developing 7 to 9 myotomes, and showing Kuffer's vesicles (Fig. 4J, K). 29 to 31 hours after the fertilization, the Kuffer’s vesicles disappeared and 11 to 12 myotomes formed (Fig. 4L). After 37 hours, lens and ear vesicles formed and the myotomes increased to 17 to 20 (Fig. 4M). After 44 hours, embryonic movements and heartbeat were observed (Fig. 4N). The hatching began 46 hours after the fertilization (Fig. 4O).
|
Each embryogenesis stage required a different time period at a different water temperature. Developing at the morula stage required an average of 5 hours at 22°C and an average of 4 hours and 30 minutes at 25°C. Blastulation required an average of 12 hours at 22°C and an average of 10 hours and 30 minutes at 25°C. Embryo formation required an average of 23 hours at 22°C and an average of 17 hours at 25°C. Hatching required an average of 46 hours at 22°C and an average of 35 hours at 25°C.
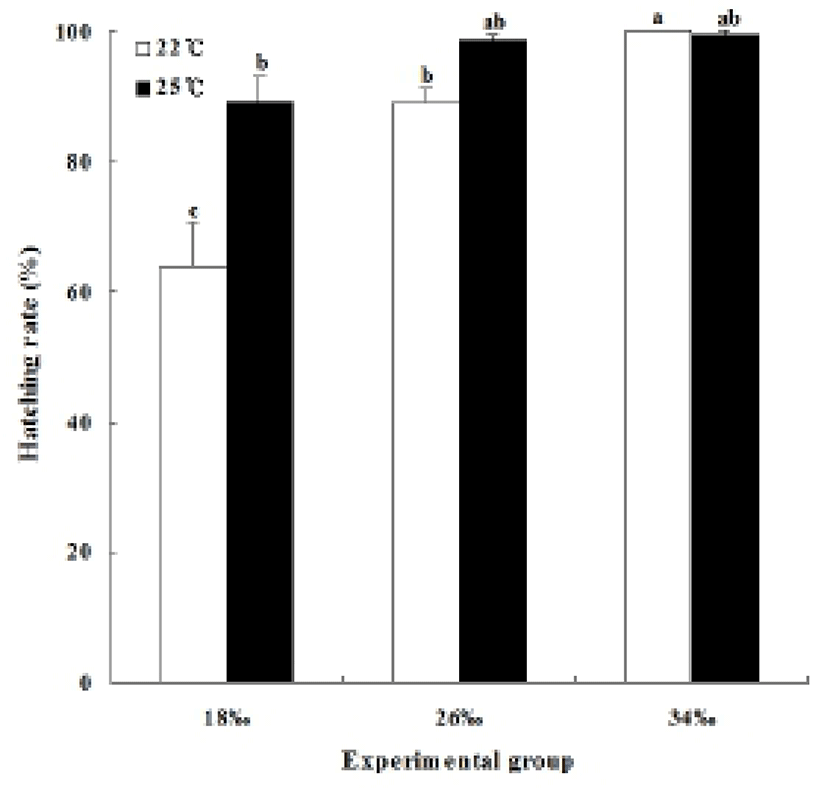
Hatching time at salinities of 18 ppt, 26 ppt, and 34 ppt showed no hatching time difference at water temperatures of 22°C and 25°C. However, the different salinities demonstrated a difference in the hatching rates at each temperature. The hatching rate at 22°C was 64.0±6.4% at the salinity of 18 ppt, 89.3±1.8% at 26 ppt, and 99.9% at 34 ppt (P<0.05). The hatching rate at 25°C had no difference between the groups of 26 ppt and 34 ppt, showing the percentage 98.7±0.7% and 99.3±0.7% respectively. However, the hatching rate was lower 89.3±3.7% at 18 ppt (P<0.05, Fig. 5) (Table 2).

|
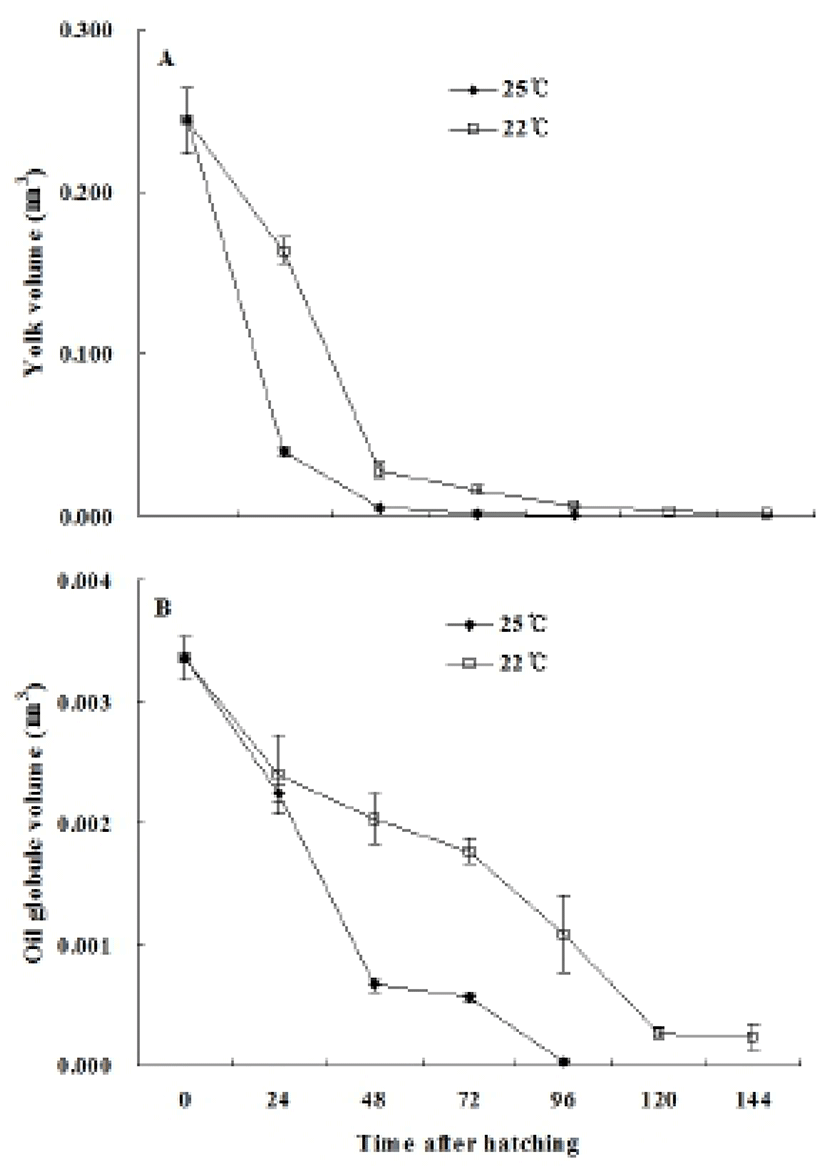
The yolk volume of the hatched larvae was 0.244± 0.020 mm3 (100%). The reared experiment groups at different temperatures showed different results. The yolk absorption was nearly complete, with 83.5% absorption, 24 hours after hatching at 25°C and 97.9% 48 hours after hatching. However, the yolk at 22°C absorbed 77.3% 24 hours after hatching, 88.2% 48 hours after hatching, and 93.1% 72 hours after hatching (Fig. 6A). The oil globules in the larvae were measured for volume at 0.0034± 0.0002 mm3 (100%). The rates of oil globule absorption at 25°C were 66.0% 24 hours after hatching, 19.7% 48 hours after hatching, and 0.9% 96 hours after hatching while the rates of oil globule absorption at 22°C were 70.5% 24 hours after hatching, 59.8% 48 hours after hatching, 31.8% 96 hours after hatching, and 7.9% 120 hours after hatching (Fig. 6B). The mouth opened in three days after hatching at 25°C, corresponding to the time period of yolk absorption and in four days after hatching at 25°C.
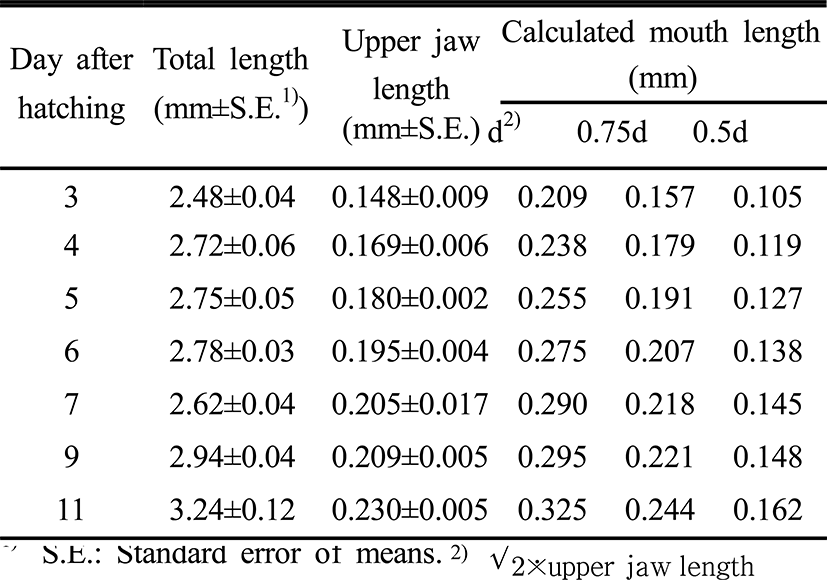
When the mouth opened as larvae three days after hatching at 25°C, the diameter (d) was 0.209 mm. The size, then, increased and was at measured 0.238 mm on the fourth day after hatching and 0.325 mm on the 11th day after hatching. The opening diameter of the hatched larvae ranged from 0.157 mm to 0.244 mm in 0.75d and 0.105 to 0.162 mm in 0.5d (Table 3).
|
The newly hatched larvae ranged from 1.75 to 2.40 mm in total length. The oil globules were located at the posterior edge of the yolk, and the anus was located a little past the body center. The melanophores were distributed on the back of mid tails and the ventral surface. The hatched larvae floated mostly near the water surface (Fig. 7A).
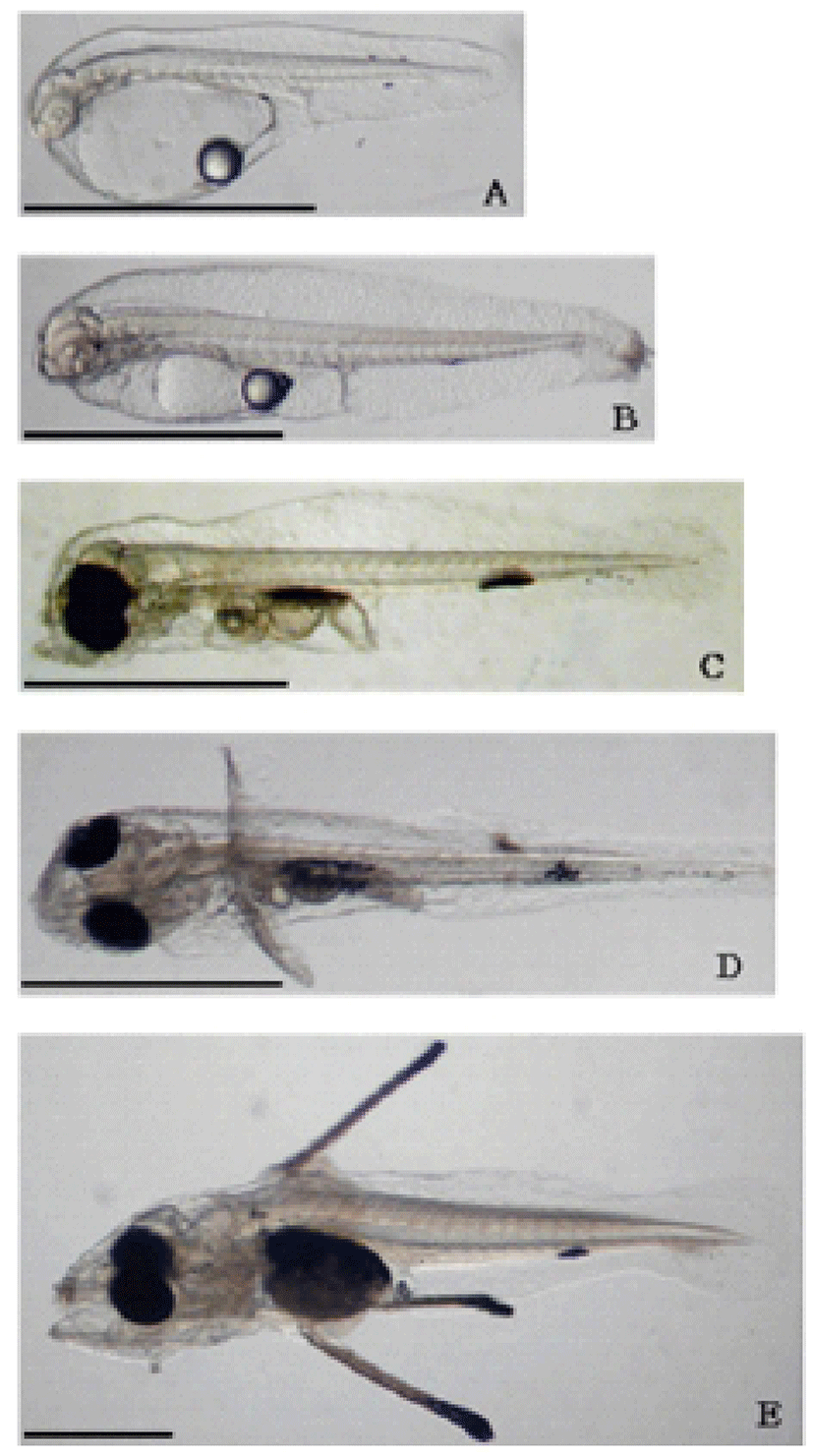
The larvae ranged from 2.46 to 2.55 mm in total length and still remained yolks and oil globules. For some of the larvae, the mouth opened 3 days after hatching. The head was formed and pectoral fins appeared. Most larvae moved to the tank bottom (Fig. 7B).
The larvae were from 2.55 to 2.72 mm in total length. Yolk and oil globules absorption was complete. The mouth and anus opened and melanophores were distributed on the lens. The dorsal fins or caudal fins were not formed yet, but the pectoral fins started to develop. The melanophores were scattered on the abdomen along the myotomes (Fig. 7D). The hatched larvae rose to the water surface and some rotifer-fed larvae were observed (Fig. 7C).
The larvae were from 2.62 to 2.94 mm in total length. The primordial fin folds started to disappear and morphological changes to form dorsal fins, caudal fins, and intestine tracts were observed. The melanophores increased on the area of abdomen and primordial fin folds (Fig. 7D).
The larvae were from 4.15 to 5.12 mm in total length. The second dorsal fins and ventral fins were formed and the layers of melanophore developed on the abdomen, forming caudal fins (Fig. 7E).
DISCUSSION
The size of fertilized eggs is one of the characteristic factors to improve larval viability, forming the total length, starvation tolerance, feeding amount, and the capability to escape from predators (Kayano, 1996). The egg sizes vary depending on the species and even in the same species, the sizes vary according to population, spawning season, and adult ages (Hamamoto et al., 1986). Even within the same subfamily Epinephelinae, location to inhabit determines the differences in the egg size. For example, E. polyphekadion, in the Red Sea, measured from 0.71 to 0.79 mm in diameter (Rasem et al., 1997), but the ones inhabiting French Polynesian averaged 0.86 mm (Aquacop et al., 1989) in diameter. E. coioides, from the Red Sea, were 0.77 mm (Hussain et al., 1975) in diameter, but the ones along the Philippines coast averaged 0.84 mm in diameter of fertilized egg (Toledo et al., 1993). The above data demonstrates that even for the same species, the larvae inhabiting a higher salinity area (42 to 43 ppt) tend to be smaller in diameter. E. akaara, inhabiting along the southern coast of Korea, measured 0.77 mm in diameter (Lee & Hur, 1997). This study on Jeju sevenband groupers showed the fertilized eggs were spherical and transparent with buoyancy. The eggs ranged from 0.76 to 0.88 mm (an average of 0.82 mm) in diameter and the oil globules were from 0.17 to 0.22 mm in diameter (an average of 0.19 mm) showing similar results from Kitajima et al. (1991). The embryogenesis and hatching rate are dependent on water temperate, salinity, light intensity, and aeration. Hatching for E. striatus requires 20 to 25 hours at water temperature ranging from 26 to 30°C (Watanabe et al., 1998). For E. polyphekadion, it takes 19 hours at 29 to 30°C (Rasem et al., 1997). The results from this study show that hatching for Jeju sevenband groupers requires 46 hours at 22°C and 35 hours at 25°C. The hatching rate showed no significant difference at different water temperatures (P>0.05) but showed low hatching rates at 22°C and 18 ppt (P<0.05).
The newly hatched larvae of E. septemfasciatus contained yolk and oil globules in the stomach and ranged from 1.75 to 2.40 mm in total length. The anus was located at the edge of the body. The yolks are the major food supplier at the early stage of embryonic development; they contain a lot of energy. Therefore, the larvae with bigger yolks can contain more energy and have more time to be fed (Bagarinao, 1986). Since the newly hatched larvae are relatively bigger than the ones from other developmental stages, the first time feeding improves viability (Hunter, 1981; Quattro & Weeks, 1991). The yolk absorption rate varies according to the rearing conditions. Epinephelinae and Siganidae, tropical marine fish species, contain relatively smaller yolks than temperate fish, but the yolk absorption occurred quicker (Bagarinao, 1986). E. akaara is known to absorb 90% of yolks at a water temperature of 25 to 29°C 24 hours after hatching and more than 90% in 64 to 80 hours after hatching (Lee & Hur, 1997). This study showed that sevenband grouper, E. septemfasciatus, consumed most of their yolks within 48 hours at the constant water temperature of 25°C but remained partial yolks at 22°C even 72 hours after hatching. The absorption of the oil globules has been reported to progress slower than the yolks (Kuo et al., 1974) due to triglyceride (Fyhn, 1989; Clyde et al., 1992). The absorption of the oil globules was done before 96 hours after hatching at the constant water temperature of 25°C but remained partially at the constant water temperature of 22°C after the mouth opened. For E. akaara, the mouth opening required 42 to 62 hours after hatching at the range of water temperature from 23 to 31°C (Lee & Hur, 1997) and 46 to 84 hours at 20 to 28°C (Kayano, 1988). The results from this study represented that sevenband groupers of mouth open required 72 to 96 hours at 22 to 25°C. The larvae were measured with the Shirota formula (1970) 0.209 mm in diameter and 0.105 mm (0.5d), larger than the minimum diameter of E. akaara larvae 0.078 mm in 0.5d for feeding (Kayano, 1988; Lee & Hur, 1997).
The dorsal fins, caudal fins, and the intestinal tracts were formed 7 to 10 days after hatching. 17 to 23 days old larvae grew from 4.15 to 5.12 mm in total length. The second dorsal fins and ventral fins were formed, demonstrating the morphological characteristics of Epinephelinae (Kitajima et al., 1991). When the larvae of sevnband grouper E. septemfasciatus grew to approximately 8.0 mm, the total length of second dorsal fins and ventral fins increased 75% and 90% respectively and then grew to adulthood. C. altivelis has a different morphological change in second dorsal fins, the total length growing 120% (Sugama & Ikenoue, 1999). E. tauvina is also noted to have differential development, growing 48% of their total length (Hussain & Higuchi, 1980). This determines egg development varies according to each species.