





INTRODUCTION
The meiotic cell cycle in mammalian oocytes proceeds up to the diplotene or dictiate stage of the first prophase and stay at this phase before LH surge (Hubbard & Terranova, 1982; Dutta et al., 2016). During growth phase of the follicle, the oocyte remains the arrested condition at prophase I. At this time it is incompetent to complete meiosis. The germinal vesicle intact oocytes in cumulus-oocyte-complex removed from mature antral follicles spontaneously progress to the metaphase II stage by culture without gonadotropin (Pincus & Enzmann, 1935; Erickson & Sorrenson, 1974). On the other hand, the oocytes enclosed in an antral follicle, do not enter meiosis even though these follicles are removed from their environment. By the adding of gonadotropin or releasing from follicle the oocyte resume meiosis. Therefore, some low molecular weight substances in the follicular fluid have been suggested as a mediator for meiotic arrest (Eppig et al., 1985; Cheon, 2004, Hwang & Cheon, 2013).
Cyclic adenosine monophosphate (cAMP) is a key signaling mediator in oocyte for meiotic prophase arrest in most species studied. Elevated cAMP concentration in the oocyte has been demonstrated to maintain meiotic arrest and a transient decrease in cAMP levels in the oocytes is thought to be a primary trigger for resumption of meiosis. cAMP works through protein kinase A (PKA)-mediated phosphorylation of maturation promoting factor (Jones, 2008). The levels of cAMP can be modulated by the activity of cAMP phosphodiesterase, PDE3A in the oocyte (Hambleton et al., 2005). On the other hand, cAMP can be transported from the surrounding cumulus cells (Norris et al., 2009) or generated by oocyte itself through the activation of heterotrimeric G protein (Gαs)-linked receptor, GPR3 or GRP12 G (Mehlmann, 2005).
In the case of cyclic GMP, it has been shown to increase in the whole ovaries in hamsters until the LH surge when cAMP rises and cGMP falls (Hubbard, 1980). cGMP works in maintaining of the meiotic arrest in oocyte and that this inhibition is dose and time dependent and expressed only in oocytes with intact cumulus (Jankowski et al., 1997; Zhang et al., 2010; Robinson et al., 2012; Egbert et al., 2014; Shuhaibar et al., 2015). It appears that the inhibitory action of cGMP on oocyte maturation is mediated by the cumulus oophorous while cAMP acts directly on the oocyte (Hubbard & Terranova, 1982). However, so far, the essentiality of cumulus cells for cGMP in oocyte is controversy. Therefore, in the present study, we investigate whether the cGMP-mediated meiotic arrest can be possible without cumulus cells during in vitro culture.
MATERIALS AND METHODS
CD1 mice were maintained on a 10 hr light and 14 hr dark cycle under standard vivarium condition and were supplied with food and water ad libitum. Immature female mice (20-24 days old) were injected with 5 IU of pregnant mares serum gonadotropin (PMSG; i.p.) to enhance multiple follicular development. PMSG primed mice are sacrificed after 46 hr and obtained the ovary. Oocytes were collected by ovarian follicular puncture with a needle under a dissecting microscope. To get cumulus free GV oocyte, the cumulus mass were removed with a fine-bore pipetted. The entire procedure did within 20 min after puncture and only the healthy oocytes and cumulus-enclosed oocytes were chosen for examination. Cumulus-oocyte complexes and oocytes were transferred into BWW medium containing 0.4% BSA and cultured.
Oocytes (8-10 oocytes/drop) were cultured in 10 µL drops of BWW in mineral oil (Sigma) or 15 hr at 37℃ in a humidified atmosphere conditioning 5% CO2. The progression of meiotic maturation was scored by observing the breakdown of germinal vesicles (GVB) and the subsequent appearance of polar bodies with an inverted microscope. Cell permeable cGMP analogs, 8-Bromoguanosine 3’,5’-cyclic monophosphate (8-Br-cGMP; 1 mM or 1 M), Rp-8-Bromo-cGMPs (1 mM or 1 M), Rp-pCPT-8-Bromo-cGMP (1 mM or 1 M) were treated into the medium and GV intact oocytes were cultured. The progression of meiotic maturation was scored by observing the breakdown of germinal vesicles (GVB) and the subsequent appearance of polar bodies with an inverted microscope.
RESULTS
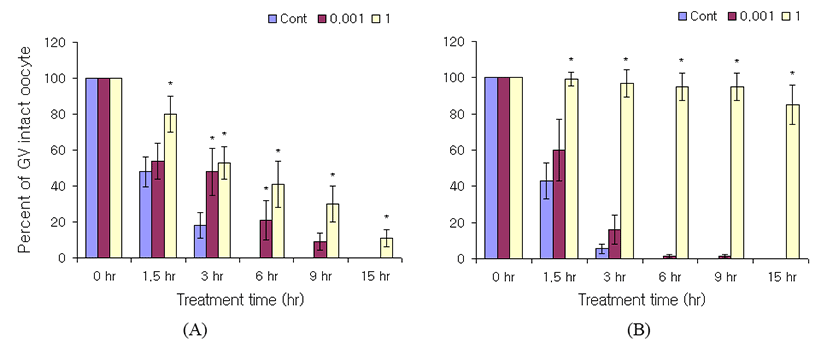
To know the difference of cGMP sensitivity to meiotic arrest by the existence of cumulus, cell permeable cGMP analog, 8-Bromoguanosine 3’,5’-cyclic monophosphate (8-Br-cGMP) was treated to GV intact oocytes as mentioned in Materials and Methods. GVB was inhibited concentration dependently by the 8-Br-cGMP in the cumulus-enclosed oocyte (Fig. 1A). In addition, it also inhibited GVB in denuded oocytes with concentration dependent manner (Fig. 1B). At 6 hr of incubation, the GV intact percentage of cumulus-enclosed oocytes was 0% at control, 21% at 1 mM 8-Br-cGMP, and 41% at 1M 8-Br-cGMP. On the other hand, in the case of denuded oocyte, the GV intact percentage was 0% at control, 1.4% at 1 mM 8-Br-cGMP, and 95% at 1 M 8-Br-cGMP. At 15 hr of incubation, the intact percentage was 0% at control, 0% at 1 mM 8-Br-cGMP, and 11% at 1 m 8-Br-cGMP. In the case of denuded oocyte, GV intact percentage was 0% both at control and 1 mM 8-Br-cGMP, and 85% at 1 M 8-Br-cGMP.
The inhibitory effect of 8-Br-cGMP was more severe in denuded oocytes than cumulus-enclosed oocytes. At 1 M 8-Br-cGMP, the GVB inhibition was significantly higher in denuded oocytes than cumulus-enclosed oocytes at all time points (Fig. 1).
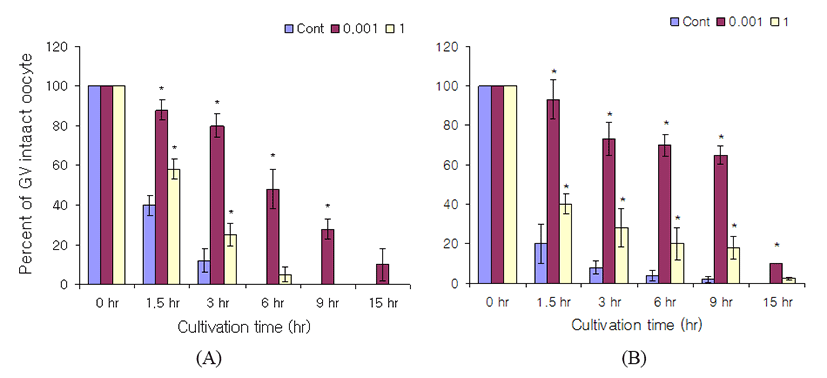
The cell permeable cGMP analog Rp-8-Br-cGMP and RP-pCPT-8-Br-cGMP but have a different function from 8-Br-cGMP were treated to the GV intact oocytes. Rp-8-Br-cGMp and Rp-pCPT-8-Br-cGMP have antagonistic effects in cGMP target cellular mediator, a cGMP-dependent protein kinase. In cumulus-enclosed oocyte, Rp-8-Br-cGMP did not block GVB (Fig. 2A). In denuded oocyte, it also did not block the GVB (Fig. 2B). However, there were no concentration dependent manners in GV arrest. At 6 hr of culture, GV intact percentages in cumulus-enclosed oocytes were 0% both at control and 1 M Rp-8-Br-cGMP, and 48% at 1 mM Rp-8-Br-cGMP. In denuded oocytes, the GV intact percentages were 4% at control, 70% at 1 mM Rp-8-Br-cGMP, and 20% at 1M Rp-8-Br-cGMP. At 15 hr after culture, the GV arrest percentages in cumulus-enclosed oocytes were 0% both at control and 1 M Rp-8-Br-cGMP, and 10% at 1 mM Rp-8-Br-cGMP. In denuded oocytes, the GV arrest percentages were 0% at control, 10 % at 1 mM Rp-8-Br-cGMP, and 2.5% at 1 M Rp-8-Br-cGMP (Fig. 2). Compared with the results of 8-Br-cGMP in denuded oocytes, the GVB blocking effects is high in 8-Br-cGMP.
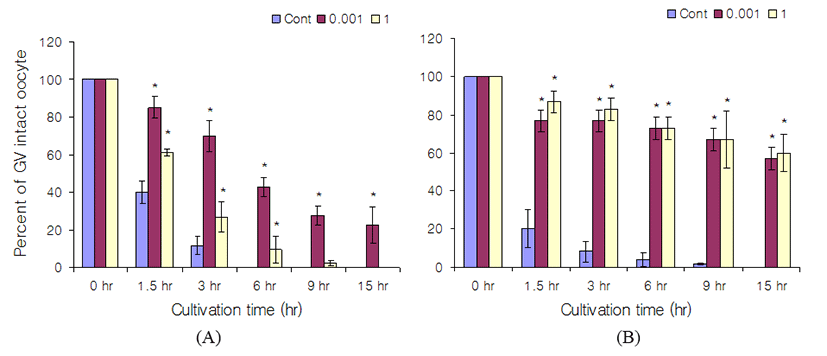
In the case of Rp-pCPT-8-Br-cGMP, its effect on GV arrest was stronger than Rp-8-Br-cGMP (Fig. 2 and 3). In cumulus-enclosed oocytes which were cultured in Rp-pCPT-8-Br-cGMP containing media, the GV arrest percenttages were 0% at control, 43% at 1 mM Rp-pCPT-8- Br-cGMP, and 9.5% at 1M Rp-pCPT-8-Br-cGMP after 6 hr of culture (Fig. 3). At 15 hr of culture, the GV arrest percentages were 0% both control and 1 M Rp-pCPT-8-Br-cGMP, and 23% at 1 mM Rp-pCPT-8-Br-cGMP. In cumulus-enclosed oocytes, the GV arrest effect is similar between Rp-8-Br-cGMP and Rp-pCPT-8-Br-cGMP. In denuded oocyte, the GV arrest percentages after 6 hr of culture were 40% at control and 73% both in 1 mM and 1 M Rp-pCPT-8-Br-cGMP. At 15 hr of culture, the GV arrest percentages were 0% at control, 57% at 1 mM Rp-pCPT-8-Br-cGMP, and 60% at 1 M Rp-pCPT-8-Br-cGMP. When compared with the results of cumulus-enclosed oocytes and denuded oocytes, the inhibitory effects of Rp-pCPT-8-Br-cGMP severe in denuded oocytes (Fig. 3).
DISCUSSION
Fertility of female is depending on the quantity and quality of follicles, and the getting a competence for development in the oocyte is critical for oncogenesis. Oocytes have to stay until ovulation at GV intact status, prophase I of meiosis. A few of cellular molecules are known to be involved in this arrest. The well-known molecules are cAMP and its metabolic materials. Cotreatment of FSH and adenosine markedly inhibits the GVB of CEO in a dose-dependent manner (Mille & Behrman, 1986). Recently, we reported about the possible roles of adenosine and adenosine receptor-mediated calcium signaling in GV arrest of oocytes enclosed with cumulus (Hwang & Cheon, 2013). It supports the role of calcium in cAMP action (Powers & Paleos, 1982; Preston et al., 1987). On the other hand, cyclic GMP also is also suggested as a mediator for cAMP regulation in oocyte. cGMP inhibit cAMP hydrolysis through PDE3A (Norris et al., 2009; Vaccari et al., 2009).
The cGMP levels in follicle are uniformly high (about 2-4 µM). Its levels are decreased to 100 nM by LH surge. The level of cGMP in oocyte is also decreased uniformly to about 100 nM and this decrease causes of meiotic restart (Shuhaibar et al., 2015). In follicle cGMP producing guanylyl cyclase natriuretic peptide receptor 2 (NPR2, guanylyl cyclase B) is present in all of the granulosa cells, but not in the oocyte (Jankowski et al., 1997; Zhang et al., 2010; Robinson et al., 2012; Egbert et al., 2014). Those are suggested that cGMP is originated from follicle cells. The cell permeable cGMP analog, 8-Br-cGMP inhibits GVB with concentration-dependent manners both in cumulus-enclosed oocytes and denuded oocyte. The effects of 8-Br-cGMP were stronger in cumulus-enclosed oocytes than denuded oocytes. Its inhibitory effects were continued until 15 hr of culture in denuded oocytes. Interestingly, the cumulus decreased the effects of cGMP on the GV arrest. Such a responsibility of oocyte to cGMP suggest that cumulus may modulate the cGMP effects on GVB arrest.
The antagonists of cGMP in cellular signaling, Rp-8-Br-cGMP and Rp-pCPT-8-Br-cGMP were showed opposite effect to 8-Br-cGMP. However, its effects were specific to the concentration of Rp-8-Br-cGMP in cumulus-enclosed oocytes but not in denuded oocytes. In denuded oocytes, the spontaneous maturation processed dependently to the concentration of Rp-8-Br-cGMP. In cumulus-enclosed oocytes, the effects of Rp-pCPT-8-Br-cGMP also had a similar pattern with the Rp-8-Br-cGMP concentration. In denuded oocytes, Rp-pCPT-8-Br-cGMP was significantly inhibited GVB in all groups compared with control, though its GVB inhibitor effect were less than 8-Br-cGMP. The responsibility to the Rp-pCPT-8-Br-cGMPthe Ministry of Education, Science and Technology and Rp-8-Br-cGMP are typically different between cumulus-enclosed oocyte and denuded oocytes. However, interestingly Rp-pCPT-8-Br-cGMP inhibited severely more than Rp-8-Br-cGMP at 1 mM but not 1 M. Such differences between antagonists for cGMP may be originated from the difference of the affinity to the target molecule. Based on them, it means that the sensitivity for GV arrest to cGMP may be modified by cumulus cells.
The cGMP transported from granulosa cells to oocyte through gap junction has been suggested as a key factor for GV arrest. Based on the results, we know that the responsibility to agonist or antagonist is modified by the cumulus. The GV arrest was maintained in denuded oocyte by 8-Br-cGMP significantly compared with Rp-8-Br-cGMP or Rp-pCPT-8-Br-cGMP. Put together, it suggests that the role of cGMP for spontaneous GVB is modulated by cumulus.