





INTRODUCTION
Pagrus major is a species of fish belonging to Pagrus genera, Sparidae families, Perciformes orders, and it is reported that there are six genera and eight species in the same genera including Acanthopagrus latus, A. schlegelii, Argyrops bleekeri, Dentex tumifrons, Evynnis cardinalis, E. japonica, Sparus sarba inhabiting Korea (Kim et al., 2005)
Overseas studies on Sparidae fish include Early life history (Seno, 1912), Growth (Oshima, 1942), Egg production and newly hatched larvae (Mito, 1957; Han & Kim, 1999), Larva and juveniles morphology development (Fukuhara, 1977; Kang et al., 2014), and Morphology development of larvae (Fukuhara, 1987), and domestic studies on Sparidae fish include Possibility of natural hybrid between species (Kang et al., 2015), The effect of water temperature on the rate of occurrence (Yoo et al., 1991), and seed production (Byun & Cho, 1982), etc. Meanwhile, larvae morphology of red seabream has been reported by some researchers such as Mito (1963) and Han & Kim (1999), but no research has been conducted on morphology from post-larvae to juvenile.
In sparidae fish, the morphology between species is similar in larva and juvenile period, so identification is difficult and research data are lacking.
In addition, when the sexual maturity is reached, there are many kinds of similar morphology, and it is difficult to identify them because they can be confused with the same species. Therefore, this study attempts to provide basic data of taxonomic study by investigating the external morphology of the larva and juveniles in developmental stages produced by artificial seedling, and identifying the relationship between sparidae fishes.
MATERIALS AND METHODS
The samples used in this study were larva and juveniles, which were hatched by naturalized spawned fertilized eggs from their mothers who were raised in an indoor round water tank (Ø 6×1 m) in May 2015 located in Sanyang-eup, Tongyeong city. Natural spawned fertilized eggs were collected, placed in a round water tank, and managed with the intensive pond system.
During the rearing period, water temperature was maintained at 22.5-23.5℃ (mean 23.0±0.5℃). As for feeding supplies, immediately after hatching to 15 days, 5-10 brackish rotifers per mL were supplied, from 16 to 25 days after hatching, 3-5 of Artemia sp. nauplius larva per mL mixed with rotifers were supplied, and from 26 to 35 days after hatching, only Artemia sp. nauplius larva were supplied. From the 40 days after hatching, extruded pellets were supplied.
For observation of larva and juveniles morphology development, five individuals were collected every 5 days, fixed in 5% neutral formalin, using a stereomicroscope (Nikon SMZ18, Japan) the length of each part was measured up to 0.01 mm, and sketched after morphology observation and photography using a digital camera installed in the microscope. The morphology development stages of larva and juveniles were classified according to Kendall et al. (1984).
RESULTS
On the 15 days after hatching, the larvae was 4.24-5.10 mm (mean 4.66±2.18 mm) in total length, and the fins of the membrane started to develop into a fan shape and the melanophore was deposited upper the alimentary canal of the abdomen and on the bladder. At this time, the stem began to form at the end of caudal fin and migrated to post-larvae (Fig. 1A).
On the 20 days after hatching, the post-larvae was 4.52-5.57 mm (mean 5.19±2.32 mm) in total length, a speckled melanophore was deposited in the head part, and four branch-shaped pigments were deposited in the anus. The 16 melanophore deposited on the caudal part were connected to each other and changed into a line shape, and the tip of the tail began to bend to the top. The anus position of this period was 41.0% towards the center of the body (Fig. 1B).
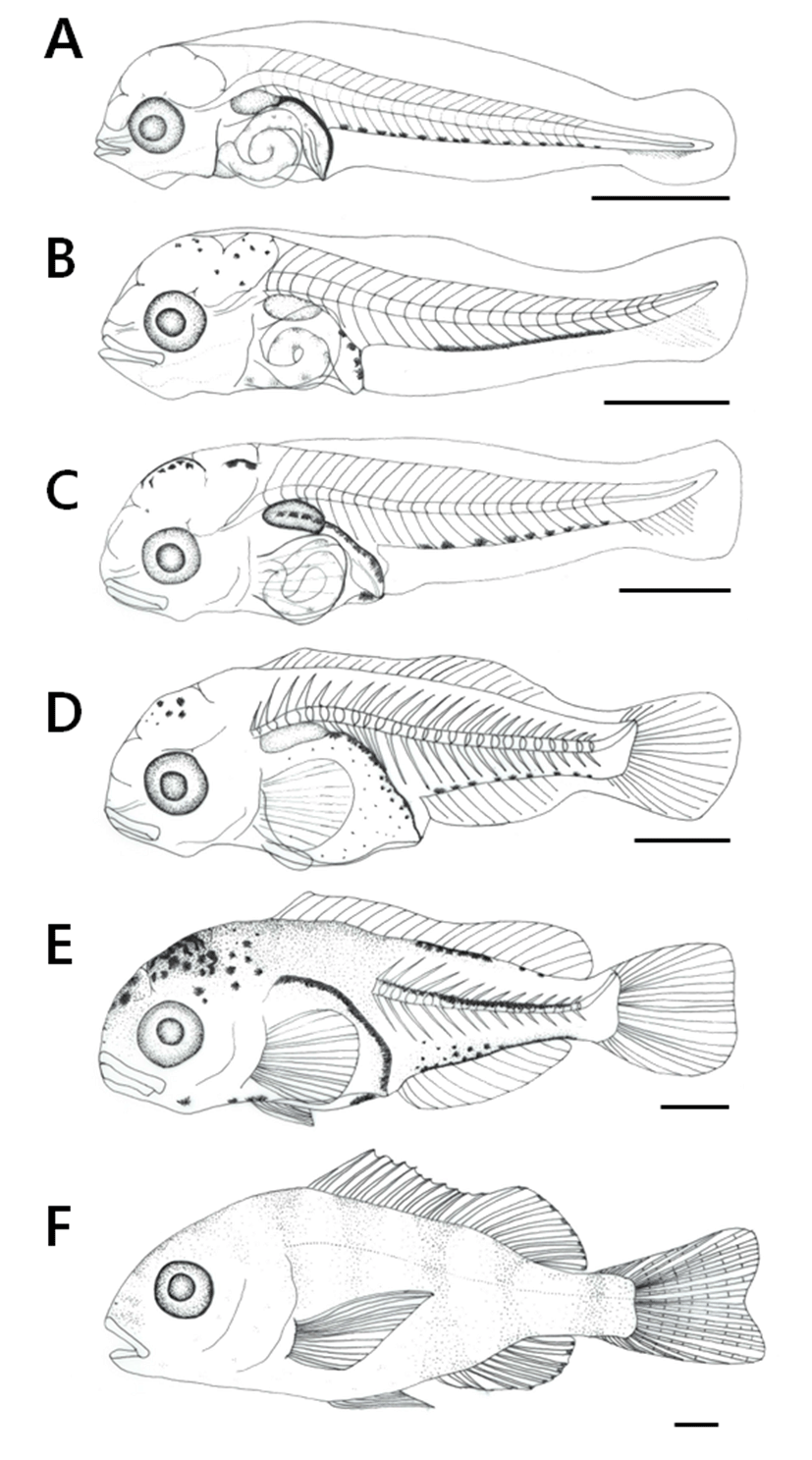
On the 25 days after hatching, the post-larvae was 4.78-6.51 mm (mean 5.76±0.39 mm) in total length, 11 rays were formed under caudal fin tibiaong, and 5 rays were formed in pectoral fin. The pigment deposited on the bladder and abdomen changed into abranch shape, and the line-shaped melanophore deposited on the caudal part again changed into nine branch shaped melanophore. The anus position of this period was 45.1% close to the center of the body (Fig. 1C).
On the 30 days after hatching, the post-larvae was 5.51-8.69 mm (mean 6.74±0.79 mm) in total length, all fins except pectoral fin were connected with membranes, but rays began to develop in each part such as dorsal, anal, and tail. 23 rays were formed in the dorsal fin, 10 rays were formed in the anal fin, and 16 rays were formed in the caudal fin along with the attached rays. At this time, ossification of the vertebrae, nerve pole, and vascular pole was made, and fins were formed in the pelvic fin. The anus position at this time was 48.8% close to the center of the body (Fig. 1D).
On the 35 days after hatching, the larvae was 6.98-12.5 mm (average 9.35±1.71 mm) in total length, a number of branch-shaped melanophore were formed on the head part, also formed on the upper and lower parts of the caudal part, and some melanophore were deposited on the underside of the head part and abdomen. The fins connected to the membrane were separated into the dorsal, tail, and anal fin, and rays were formed in the pelvic fin. The number of partial fins increased to 12 spinous rays and 10 soft rays in the dorsal fin, 3 spinous rays and 8 soft rays in the anal fin, which reached the number of fin rays coincided with that of the mother and proceeded to juvenile (Fig. 1E).
On the 40 days after hatching, the juvenile was 11.3-18.1 mm (mean 14.9±1.53 mm) in total length, and the
side line was clearly formed along the side of the body. The translucent part of the body was covered with scales and developed opaque. The melanophore, which was deposited on the body, was formed with 6 segment sand the pectoral fin tip was long developed and similar to the morphology of the mother. The anus position at this time was 57.8%, located beyond the center of the body (Fig. 1F).
Larvae and juvenile growth of red seabream is shown in Fig. 2. Immediately after hatching, the mouth was not open and up to the 3 days after hatching when the yolk was completely absorbed under the condition of no feeding activity, the growth was gentle to an average total length of 2.78 mm. Since then, steady growth was shown and at an average total length of 5.76 mm on the 25 days after hatching, Artemia sp. nauplius larva alone began to be supplied, and the growth accelerated in a short period of time up to an average total length of 6.74 mm within 30 days after hatching. From 31 to 34 days after hatching, fast growth was observed, and from the average total length of 9.35 mm within 35-36 days after hatching to the average total length of 14.9 mm within 40 days, the juvenile showed rapid growth. During this period, the supply of food organisms was stopped and extruded pellets were alternatively supplied alone, and in 40 days after hatching, the growth was again gentle.
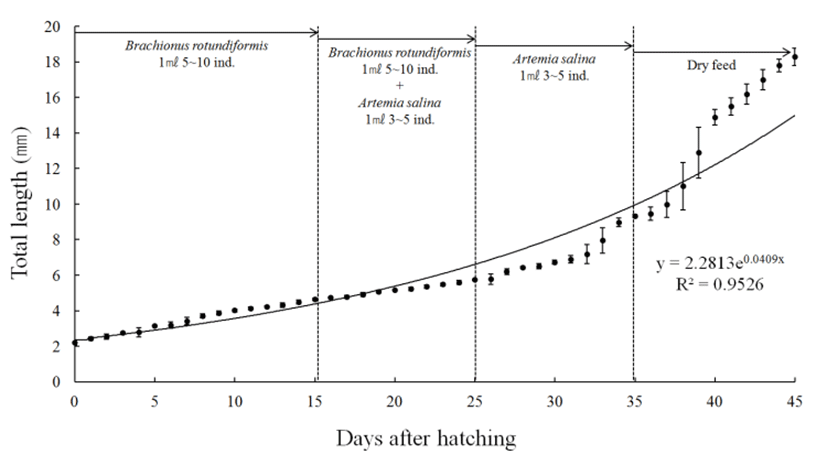
Larva and juveniles of red seabream showed fast growth when the type of food supply was changed, and gentle growth rate was shown after 3 to 5 days of the supply.
DISCUSSION
In order to establish marine fish production technology and identify natural resources in nature, it is necessary to know the initial development of the target species. In particular, it is important to observe the morphological features of eggs and larva and juveniles by artificial insemination in the indoor environment, and to compare them with other species, because it is difficult to identify the species in natural eggs and larva and juveniles collected from the nature (Byun et al., 2009).
Based on the results of this study, we compared the relative relationships among the relative species. From the results of comparing total lengths when moved to post-larvae, red seabream entering post-larvae stage was 4.66 mm in average total length, and according to the results of Han & Kim (1999), their average total length was 3.28 mm when entering post-larvae stage, where both studies showed differences. It is thought that water temperature, food, and breeding environment are related to the growth rate of larvae, and to identify this, many further related researches will be needed in future. In contrast to the total length of other fish in the post-larvae period, the average total length of O. fasciatus (Koh & Kim, 1992) was 4.49 mm and that of A. schlegelii (Kang et al., 2014) was 4.45 mm, while O. punctatus (Park et al., 2015) was longer at 8.83 mm.
During the transition to the post-larvae period, 15 melanophores were deposited in a tree branch shape from the lower abdomen towards the tail when the average total length of red seabream was 4.66 mm, and melanophores on the upper digestive tract were darkly pigmented.
Melanophores increased in O. fasciatus (Koh & Kim, 1992) from occiput when its average total length was 4.49 mm, and were deposited in a speckled arrangement in the upper maxillary, opercular, and central parts of the body in O. punctatus (Park et al., 2015) when its average length was 8.83 mm. In A. schlegelii (Kang et al., 2014), 15 melanophores were deposited on the upper and lower parts of the digestive tract and lower abdomen when its average total length was 4.45 mm. The total length and the number of deposited melanophores were similar in the lower abdomens of red seabream and A. schlegelii, which may lead to confusion. However, unlike A. schlegelii, melanophores were not deposited in the lower part of the digestive tract in red seabream, and this thus serves as a point of differentiation between the two (Table 2).
| Species | Days untillpostlarvae (day) | Postlarvae size (mm) | Days untilljuvenile (day) | Juveniles size (mm) | References |
|---|---|---|---|---|---|
| Pagrus major | 15 | 4.66(4.24-5.10) | 35 | 9.35(6.98-12.5) | Present study |
| - | 3.28 | - | - | Han & Kim, 1999 | |
| Acanthopagrus schlegelii | 5-6 | 4.45(4.32-4.66) | 30-32 | 15.8(15.4-16.1) | Kang et al., 2014 |
| Oplegnathus fasciatus | 14 | 4.49(4.13-4.67) | 28 | 10.4(9.96-10.2) | Koh & Kim, 1992 |
| Oplegnathus punctatus | 18 | 8.83(8.71-8.94) | 35 | 23.2(21.5-24.8) | Park et al., 2015 |
| Species | Melanophore distribution | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Postlarvae( mm ) | Otocyst | Dorsal | Ventral | Center of tail | Front of eye | Oil globule | Back of head | Abdomen | |
| Pagrus major (Present study) | 4.66 | - | - | + | + | - | - | - | + |
| Acanthopagrus schlegelii (Kang et al., 2014) | 4.45 | - | - | + | - | - | - | - | + |
| Oplegnathus punctatus (Park et al., 2015) | 8.83 | - | - | + | + | - | - | + | - |
| Oplegnathus fasciatus (Ko & Kim, 1992) | 4.49 | - | - | - | - | - | - | + | - |
When comparing the period of caudal notochord bending, the caudal notochord of red seabream began to bend at 30 days after hatching with an average total length of 6.74 mm. Park et al. (2015) showed that the caudal notochord of O. punctatus began to bend at 15 days after hatching with an average total length of 7.24 mm, Koh & Kim (1992) showed that the caudal notochord of O. fasciatus began to bend at 24 days after hatching with an average total length of 8.90 mm, and Kang et al. (2014) showed that the caudal notochord of A. schlegelii began to bend at 16-18 days after hatching with an average total length of 7.02 mm, showing that there are various differences according to the time and the size of the total length in every species.
In comparison with the time of entering juvenile stage, at 35 days after hatching when the average total length was 9.35 mm, fin rays of red seabream were formed and finally the larvae entered juvenile stage, and the average total length of O. punctatus (Park et al., 2015) was 23.2 mm at 35 days after hatching, which is longer than that of red seabream. The average total length of O. fasciatus (Koh & Kim, 1992) was 16.7 mm at 37 days after hatching, and the average total length of A. schlegelii (Kang et al., 2014) was 15.8 mm at 30-32 days after hatching, which were shorter than that of red seabream, demonstrating differences.
To summarize the results of this study, the total length of post-larvae red seabream was similar to that of other closely related species, but red seabream exhibited differences in the distributional location of melanophores, and thus could be morphologically classified. The period in which the bending of the cordate terminal and the transition to the juvenile stage occurred differed between species, since this occurred in red seabream when its total length was the shortest of all species examined (Table 1).
Studies on the larva and juveniles morphology of sparidae fish inhabiting Korea have not been studied sufficiently until now. In order to establish the taxonomic study data, it is necessary to continue research on the early life history through securing the mother, breeding larva and juveniles.
In addition, bone abnormalities occur in larvae and juveniles in red seabream which is the main cause of early mass mortality and affects yield. Therefore, further studies on the skeletal development of larva and juveniles should be performed in order to identify the exact causes.