
INTRODUCTION
The marine medaka, Oryzias dancena, is a truly euryhaline teleost fish, having a great capacity for hypo and hyper-osmoregulation. Most of its physiological attributes are similar across a wide spectrum of salinities, ranging from fresh water to normal seawater (Inoue & Takei, 2003; Kang et al., 2008; Cho et al., 2010). Therefore, much attention has been directed at extending the utility of functional transgenic marine medaka strains for ornamental purposes, because they can be used at most naturally occurring salinities (Cho et al., 2011). In addition, in a recent study of transgenic marine medaka containing the myosin light chain-2 (mlc2f) promoter, the expression of a vivid red fluorescent color in their fast skeletal muscles suggested great potential for these as novel ornamental fish for both freshwater and seawater aquaria (Cho et al., 2012). Triploidization is a technique used to generate sterile aquatic animals by taking advantage of the incompatibility in pairing the three homologous chromosomes during meiosis I (Don & Avtalion, 1986). This technique has also been used to enhance the productivity of several fish species because of its assumed ability to increase yield by channeling the energy required from gonadal development to somatic growth (Tave, 1993). More importantly, it generates fish that are unable to breed and contribute to the local gene pool if they were to accidentally escape from confinement. By conferring sterility of exotic fish for a limited purpose, triploidy can serve as an effective method for reducing or eliminating the environmental risks of genetically modified organisms (Kim et al., 1994).
Numerous studies concerning the growth rates of triploid fish have been published. Growth rates tend to slow or cease in maturing fish, and it is during the later stages of sexual maturation that a growth advantage of triploid fish over diploids is most likely to be observed (Hatanaka et al., 1991; Kobayashi, 1992; Benfey, 1999). In species that survive spawning, diploids frequently exhibit compensatory growth and may overcome any disadvantage (Benfey, 1999). Therefore, a growth advantage of triploids is most likely to be seen in species where the diploids have high or complete mortality associated with sexual maturation and spawning, as in ayu (Plecoglossus altivelis) and twicespawned rainbow trout (Oncorhynchus mykiss) (Hatanaka et al., 1991; Kobayashi, 1992). Thyroid stimulating hormone (TSH) and thyroxine (T4) play indispensable roles during the embryonic and larval periods of fish development (Khalil et al., 2011). Because of the potentiating effects of TSH and T4 on fish larval growth and survival, it is important to investigate whether application of exogenous thyroid stimulating hormone and thyroxine to female brood fish results in better growth and survival of larvae (Khalil et al., 2011).
No previous study of the marine medaka has included a comparative analysis of the diploid and triploid fish. Therefore, we undertook a comparative analysis of the comparative study of growth and gonad maturation in diploid and triploid marine medaka, Oryzias dancena. So, the objectives of this study were to evaluate the reproductive characteristics of the triploid form of this species.
MATERIALS AND METHODS
The specimens of marine medaka, Oryzias dancena used in this study were from a laboratory stock maintained at the Institute of Marine Living Modified Organisms (iMLMO), Pukyong National University, Busan, Korea. The general maintenance of the experimental fish was according to the method of Song et al. (2009). Breeding occurred in brackish water (5 psu), as described by Cho et al. (2010). The breeding conditions included a temperature of 25 ± 1°C and a 16 h light:8 h dark cycle. Triploid and diploid specimens were fed brine shrimp (Artemia nauplii; INVE, Salt Lake City, Utah, USA) and micro-particle feed (150–500 µm diameter; Ewha Oil Co., Busan, Korea).
Fertilized eggs were obtained each day by mating male and female broodfish in a glass tank containing 30 L of well-aerated water. The eggs were collected from females immediately following fertilization. At 2 min after fertilization the fertilized eggs were subjected to a cold shock treatment at 0°C for 45 min. Following treatment the eggs were placed in a 25°C incubator until they hatched. The hatching success and the incidence of abnormal larvae were assessed based on stereoscopic microscope (C-DS; Nikon Co., Tokyo, Japan) examination of at least 23 eggs per group. The values for these parameters in the experimental treatments were expressed as percentages of treated eggs. After 60 days, triploid marine medaka were distinguished by flowcytometry and chromosome analysis.
Over 12 months the standard length, body weight, condition factor, and gonadosomatic index (GSI) for each individual 120 were measured to investigate the growth and maturation of diploid and triploid fish. The condition factor was determined using the equation: condition factor = (body weight × 100)/(body length)3, and the GSI was determined using the equation: GSI = (gonad weight/body weight) × 100. Sex hormone and growth hormone were analyzed through the plasma of each ploidy group at 120 days after hatching. The estradiol-17ß and testosterone concentrations were measured using fluorophotometry method while 48 hrs (i-Chroma, Sun Kyung Medical, Korea). To measure the concentrations of estradiol-17ß and testosterone, draw a 75 uL of serum and add it to the detector buffer. Next, mix a specimen with buffer by voltex mixer and leave to the specimen at room temperature for 15 min, finally, insert it to the i-Chroma reader.
To investigate changes in growth hormone levels, the concentrations of thyroid stimulating hormone (TSH) and thyroxine (T4) in samples of each group were measured using fluorophotometry during the 12 months following hatching. To measure the concentrations of TSH and T4, draw a 10 µL of serum and add it to the detector buffer. Next, mix a specimen with buffer by voltex mixer and leave to the specimen at room temperature for 10 min, finally, insert it to the i-Chroma reader.
The gonad morphology was observed in one-year-old diploid and triploid males and females. The gonads were surgically removed and fixed in buffered 10% formaldehyde solution for histological analysis, following morphological analysis. Conventional histological techniques were used to assess gonad development, including embedding in paraffin wax, sectioning to 6 µm thickness, and staining using Mayer’s hematoxylin and eosin.
The standard length, body weight, condition factor, and GSI were assessed using ANOVA followed by Duncan’s multiple range test at the significance level P=0.05.
RESULTS
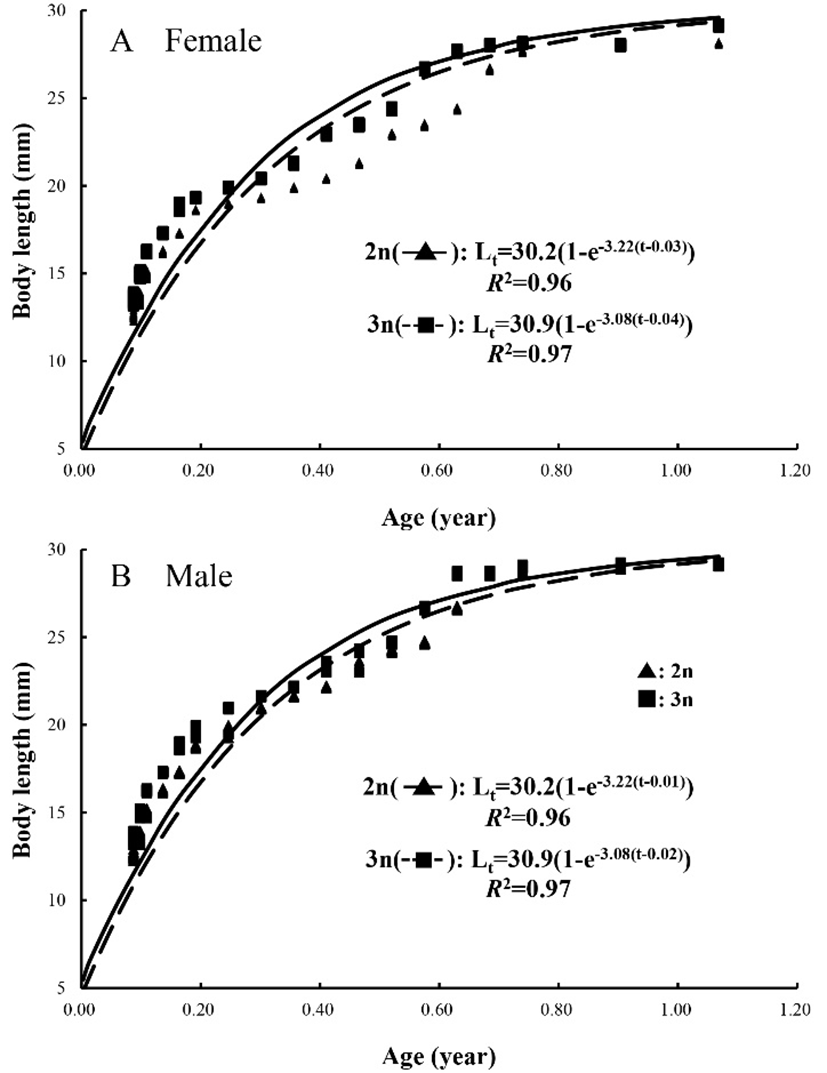
The von Bertalanffy growth parameters for diploid and triploid marine medaka, Oryzias dancena estimated using the non-linear regression method, are shown in Fig. 1. The von Bertalanffy growth equations were Lt=30.2(1–e–3.22(t–0.01)) and Lt=30.9(1–e–3.08(t–0.02)) for the diploid and triploid females, respectively. The growth coefficients (K) for the diploid and triploid females were estimated to be 3.22/year and 3.08/year, respectively, their asymptotic maximum length (L∞) was estimated to be 30.2 mm and 30.9 mm, respectively, and the theoretical age at zero length (t0) was estimated to be –0.03 and –0.04, respectively. The von Bertalanffy growth equations were Lt=30.2(1–e–3.22(t–0.01)) and Lt=30.9(1–e–3.08(t–0.02)) for the diploid and triploid males, respectively. The K and L∞ values for the diploid and triploid males were similar to those of the females. The t0 values differed between the diploid and triploid males (–0.01 and –0.02, respectively), and was greater than that for the females. For each measured parameter, significant differences were found between the diploids and triploids, and the males and females (P<0.05).
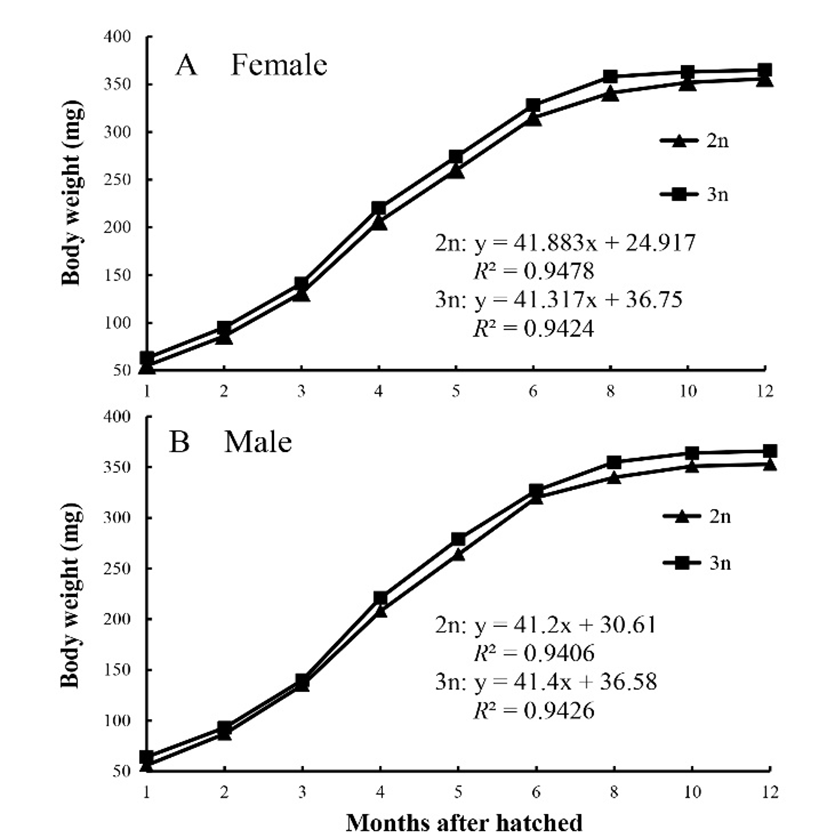
The change in body weight for diploid and triploid marine medaka is shown in Fig. 2. During the experimental period the growth patterns for the diploids and triploids (female and male groups) were similar; all fish grew rapidly from 3 to 4 months of age, and grew slowly from 8 to 12 months of age. For both males and females the triploids were larger than their diploid counterparts during the experimental period. Among both diploid and triploid fish the body weight of males was greater than that of females. Fish condition was affected by ploidy, but not by gender v (Table 1; P<0.05), and the condition of all groups decreased rapidly from 1 to 2 months of age. Up to 4 months of age the condition factor for both the male and female diploids was significantly higher than for their triploid counterparts. However, after 4 months of age the condition factor for both male and female groups was not significantly different between the diploid and triploid fish. During the experimental period, the condition factor within the diploid and triploid groups did not differ significantly between females and males.
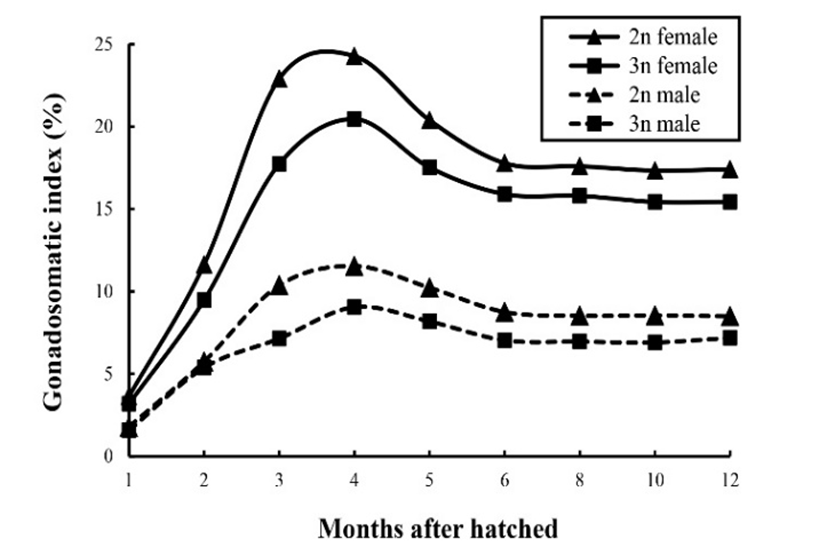
Changes in the GSI for diploid and triploid marine medaka are shown in Fig. 3. The GSI for triploid females and males was lower than that for their diploid counterparts, and within each ploidy group the GSI for females was higher than that for males. Changes in the GSI followed a similar pattern in all groups. The GSI of triploid females increased from 3.17% at 1 month following hatching to 20.45% at 4 months (P<0.05), and decreased to 15.43% at 12 months. The GSI of triploid males increased from 1.56% at 1 month following hatching to 9.05% at 4 months (P<0.05), and decreased to 7.18% at 12 months. For all groups the GSI was highest at 4 months, and decreased from 4 to 12 months. Morphological and histological analyses of the gonads of one-year-old fish showed that the triploid genotype caused significant depression of gonad development in marine medaka. The ovaries of the diploid females were well developed, with fully yolk-laden eggs, whereas those of the triploid females were poorly developed and significantly smaller than in the diploid fish (Fig. 4a and 4b). In contrast, there was no clear visual difference in the testicular morphology of diploid and triploid males, although the testes of the triploid fish were slightly smaller, suggesting that the GSI of the triploid males differed from that of their diploid counterparts (Fig. 4c and 4d).

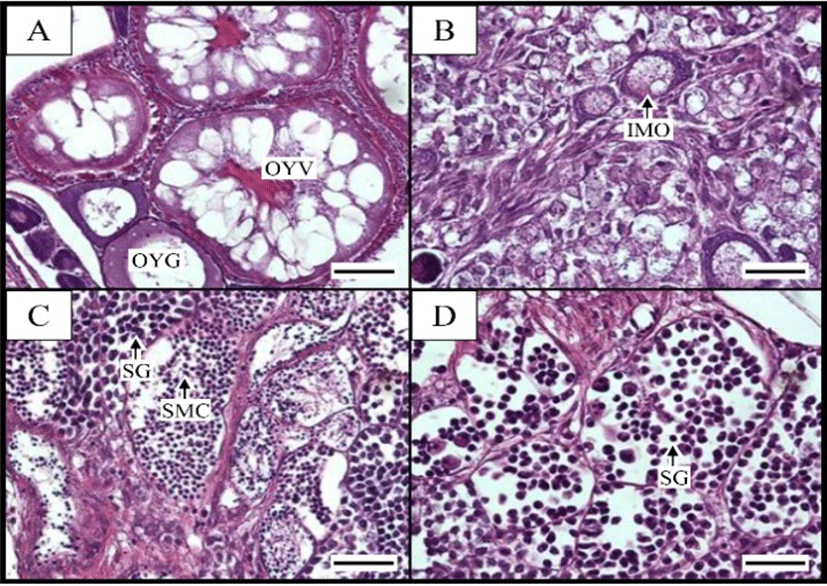
Histological analysis of the gonads showed clear differences between the diploid and triploid females. Unlike the diploid fish, which had highly developed ovaries filled with mature yolk-containing oocytes, the ovaries of the triploid females contained a considerable number of oogonia, with very few oocytes at the chromatin–nucleolus stage in the mesenchymal tissue (Fig. 5a and 5b). In addition, the triploid males showed depressed gonad development compared with their diploid counterparts, even though the gross morphology of the diploid and triploid testes was not significantly different. Normal spermatids and sperm were present in the testes of the diploid fish, whereas the testes of the triploid males had few spermatids or no sperm (Fig. 5c and 5d).
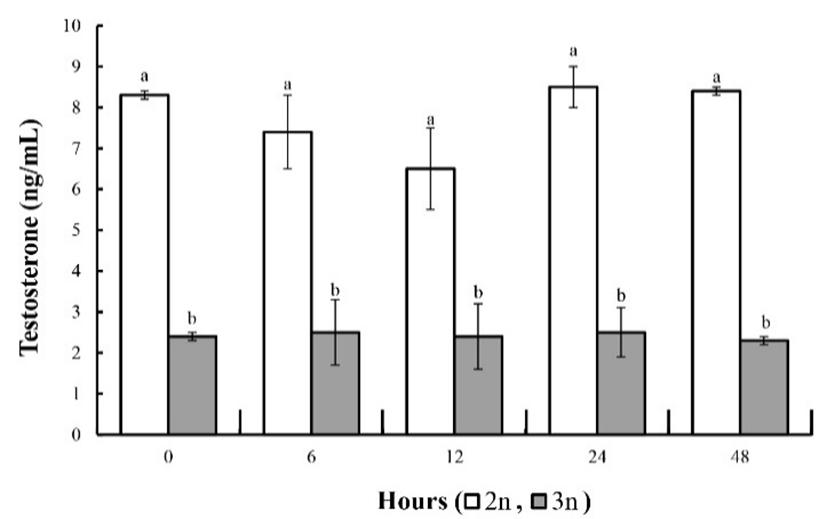
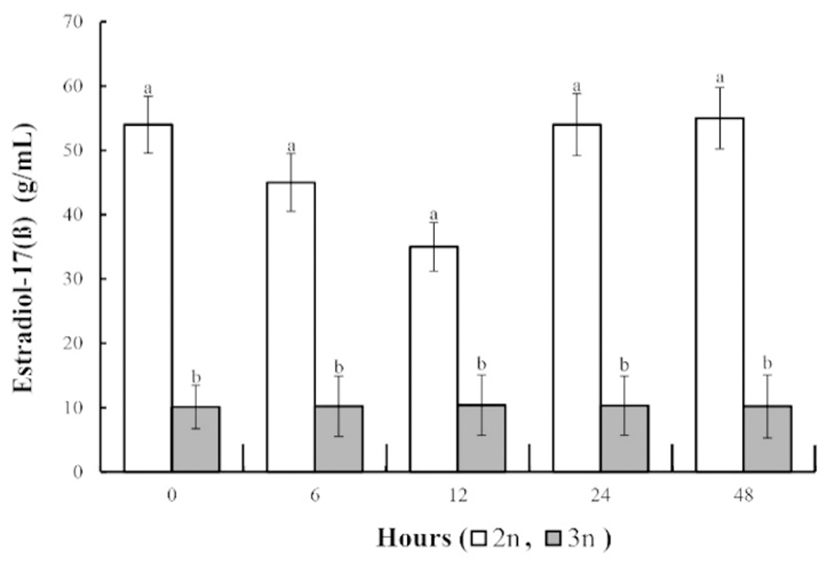
The concentrations of thyroid stimulating hormone and thyroxine were affected by the ploidy of fish, but not by their gender (Table 2; P<0.05). The concentration of thyroid stimulating hormone and thyroxine in all groups increased slowly during the experimental period. In both the diploid and triploid groups the concentrations of these two hormones were lower in the females than in the males, but for both genders were higher in triploid fish than in the diploids. As shown in Fig. 6, the testosterone concentration in the diploid fish gradually decreased after the experiment. At 24 h, values of testosterone restored initial value, and at 48 h the testosterone concentration was similar to that at 0 and 24 h. However, the triploids had lower testosterone concentrations than the diploids because of their lower GSI (P<0.05; Fig. 6). The estradiol-17ß concentration in the diploid fish showed a tendency to gradually decrease from 6 to 12 h (Fig. 7). However, at 24 and 48 h the estradiol-17ß concentration had returned to the level at 0 h. In the triploids the estradiol-17ß concentration did not change, and the concentrations of testosterone and estradiol-17ß in the induced triploids was significantly less than in the diploids (P<0.05; Fig. 10).
DISCUSSION
The triploid marine medaka, Oryzias dancena grew more rapidly than their diploid counterparts (P<0.05), which is consistent with the results of previous studies. Nam et al. (2001) reported that triploid mud loach had a growth rate 22–25 times that of diploid fish. In this study the triploid marine medaka were larger than the diploid form, but giantism of the triploid was not responsible. Kim et al. (2001) & Seol et al. (2008) suggested that the absence of giantism in triploids is because of their smaller cell number. In the present study the male marine medaka (diploid and triploid) were larger than the females (P<0.05). Female fish are typically larger than male fish of the same age, although in some species the reverse is true; examples include the gudgeon, Gobio gobio (Mann, 1980) & the filefish, Brachaluteres ulvarum (Akagawa et al., 1995). The reasons for these size differences are unclear (Katano, 1998). It has been suggested in several studies that the evolution of a larger body size in males probably results from male–male competition associated with a polygynous mating systems (Katano, 1998; Kim et al., 2008). Therefore, exploring the nature and extent of sexual dimorphism can extend our understanding of social structure and adaptation, as well as species identification. Over 1 month we found that a rapid increase in length caused a reduction in condition, as measured using the condition factor, and the condition factor for triploids was lower than that for the diploids. Unfortunately, no previous studies have reported such a rapid decrease in condition, or differences in the condition factor between diploid and triploid marine medaka or other fish species, so our observations remain unexplained.
Morphological and histological analyses of triploid gonads probably causes sterility in this species. The odd chromosome number induced during triploidization precludes homologous chromosomal pairing during meiosis I, which leads to inefficient gamete differentiation and consequently confers sterility on the triploid fish (Zhang et al., 2005). Overall, most of our observations of the gonads in this study were similar to those previously reported for other triploid fish: smaller gonads, significantly delayed gonadal development, and more pronounced sterility in females (Kim et al., 1994; Felip et al., 1999; Tiwary et al., 2000; Feindel et al., 2011). In a recent study Cal et al. (2010) reported that the induction of triploidy lowered the estradiol-17ß concentration and changed its internal secretion, resulting in oogenesis disorders. However, numerous previous studies have also claimed that triploid fish, especially old triploid males, may have a functional but reduced capacity to reproduce, despite their sterile-like gonadal development (Benfey, 1999). The artificial insemination of normal haploid eggs of tench (Tinca tinca) using sperm from triploid fish has been reported to produce some abnormal larvae (Linhart et al., 2006). In a recent study, Karami et al. (2011) reported that treatment of triploid African catfish (Clarias gariepinus) with the ovulation/ spermiating agent Ovaprim induced advanced vitellogenin sequestration in triploid oocytes, and promoted the fertilization capacity of milt from triploid males.
Sex hormones, including testosterone and estradiol-17ß, are commonly found in induced triploids because their testes and ovaries don't mature (Kim et al., 1994; Park & Kim, 1994). Unlike the normal gonadal maturation of diploids, in induced triploids the gonads are immature on formal and histological (Lincoln & Scott, 1984; Kim et al., 1994), and induced triploids showed lower concentrations of sex hormones because of decreased hormone secretion resulting from immaturity (Lincoln, 1981; Lincoln & Scott, 1984). The testes of diploids had normal spermatids and spermatozoa, while few were seen in the induced triploids, and the ovaries of diploids had many well developed oocytes, while those of the induced triploids exhibited oogonia (Kim et al., 1994; Park & Kim, 1994; Kim et al., 2001).
The concentrations of thyroid stimulating hormone (TSH) and thyroxine (T4) were found to be higher in the induced triploid fish over 1 year (P<0.05). In both the diploid and triploid groups the concentrations of TSH and T4 were higher in males than in females. In previous studies the initial survival of freshwater fish, transformation, and the effects of TSH and T4 on development and growth have been observed (Lam & Sharma, 1985; Weatherley & Gill, 1987). Administration of growth hormone caused a reduction in the weight to length ratio (condition factor) in diploids but not in triploids, and caused triploids to deplete lipid energy stores more rapidly (McLean et al., 1991). In our experiment the induced triploids were expected to show better growth compared with the diploids, and the condition factor for the latter was expected to decrease faster. As no comparable studies have investigated differences in TSH and T4 between diploid and triploid in marine medaka or other fish species, no explanation for our observations is available. Although induced triploids are infertile, their production is economically feasible because they grow faster than their diploid counterparts. Further studies will be necessary to achieve stable production of triploids. In particular, longer-term observations of the potential for maturation in older triploid marine medaka should extend our understanding of their reproductive capacity.