
INTRODUCTION
Liobagrus obesus is a fish belonging to Siluriformes Amblycipitidae Liobagrus, and it is known to be distributed in the Geumgang River and the Ungcheon River, Mangyeong River and upper stream of Yeongsan River. Amblycipitidae fish are commonly characterized by four pairs of barbells and a thorn on the pectoral fin, and there are three species known in Korea including L. andersoni and L. mediadiposalis as well as L. obesus (Kim et al., 2005).
L. obesus and L. andersoni have been in confusion due to their similarities in shape, but through taxonomic studies, these are divided by the number of pectoral fins and the number of chromosomes according to the distribution area (Son & Lee, 1989). In addition, L. obesus is a species native to Korea and has very limited habitats and fewer populations, and is designated and protected by the Ministry of Environment in 1998 as an endangered species of wild fauna and flora (NBC, 2018).
Studies of Liobagrus include the early life history (Seo et al., 2006), spawning behavior and early life history of L. mediadiposalis (Choi et al., 2008), feeding habits of L. obesus (Son & Byeon, 2004), characteristics of natural spawning ground (Kim et al., 2012), feeding habits of L. andersoni (Yoon et al., 2007), metrological trait characteristic (Park et al., 2006) and karyotype analysis (Son & Lee, 1989), etc. However, there has been no study of skeletal development for taxonomic studies.
Studies on the skeletal development of fishes are important for the identification of species as well as skeletal characteristics of fishes, so detailed and systematic studies are needed (Koumoundouros et al., 1997a,b). Also, the stage of morphogenesis of the adolescent period helps to understand the systematic basic trait expression, functional development tendency and preference for the environment, and research in this field is diverse (Mook, 1977; Potthoff et al., 1988; Fukuhara, 1992; Faustino & Power, 1999; Koumoundouros et al., 1999; Liu, 2001; Sfakianakis et al., 2004).
In particular, three species of Liobagrus fish have been attracting attention as an important research area due to regional differences in form. So far, the research on the internal skeleton of Liobagrus fish has been lacking, and this study examines larvae skeletal development of L. obesus to use the results as a basic data for taxonomic research and artificial proliferation.
MATERIALS AND METHODS
The brood stork fish used in this study were secured during the research on the preservation, restoration and propagation technology development of the endangered species of Korean specialty fish, which is the next generation core environment technology development project of Ministry of Environment (Project No. 652-041-006), and the research was conducted in May 2004, in the Chogangcheon Stream located in Yeongdong-gun, Chungnam Province. The embryos of L. obesus were collected under the rocks where the flow speed was relatively gentle. Collected embryos were transferred to the laboratory and housed in a rectangular glass water bath (35×50×30 cm). The water temperature was maintained at 19.5-24.9°C (average 22.2± 3.81°C) by using a sponge filter.
Larvae were kept in a circulatory system in the fertilized egg management tank. The temperature of the rearing water remained the same as the embryos. Feeding of Artemia sp. nauplius larvae supplied at 5-10 per mL was carried out from the 10th day to the 16th day after hatching and absorbing egg yolk. Sludge worms were supplied from 16th to 24th day after hatching. From 24th day after hatching, frozen blood worms were supplied. For skeletal developmental observation, five individuals were collected every day from immediately after hatching and fixed to 10% neutral formalin. Skeletal staining was performed by double staining of Kawamura & Hosoya (1991). The stained larvae were stored in 50% Glycerin. Sections of each skeleton were sketched using a universal projector (Nikon JP V-12B, Japan) and a stereomicroscope (Nikon NM-40, Japan) to sketch the developmental process. The names of each site followed Kang et al. (2012) and Park et al. (2015).
RESULTS
For observation of skeletal development of L. obesus larvae, it was divided into cranium, visceral skeleton, shoulder girdle bone, pelvic girdle bone and vertebra.
The development of cranium and visceral skeleton of L. obesus larvae is shown in Table 1. Shortly after hatching, the pre-larvae showed a linear parasphenoid ossification in the cranium with an overall length of 7.52-7.69 (mean 7.60±0.12) mm. In the jaw bone, the dentary supporting the lower jaw and the maxillary supporting the upper jaw were ossified (Fig. 1A).
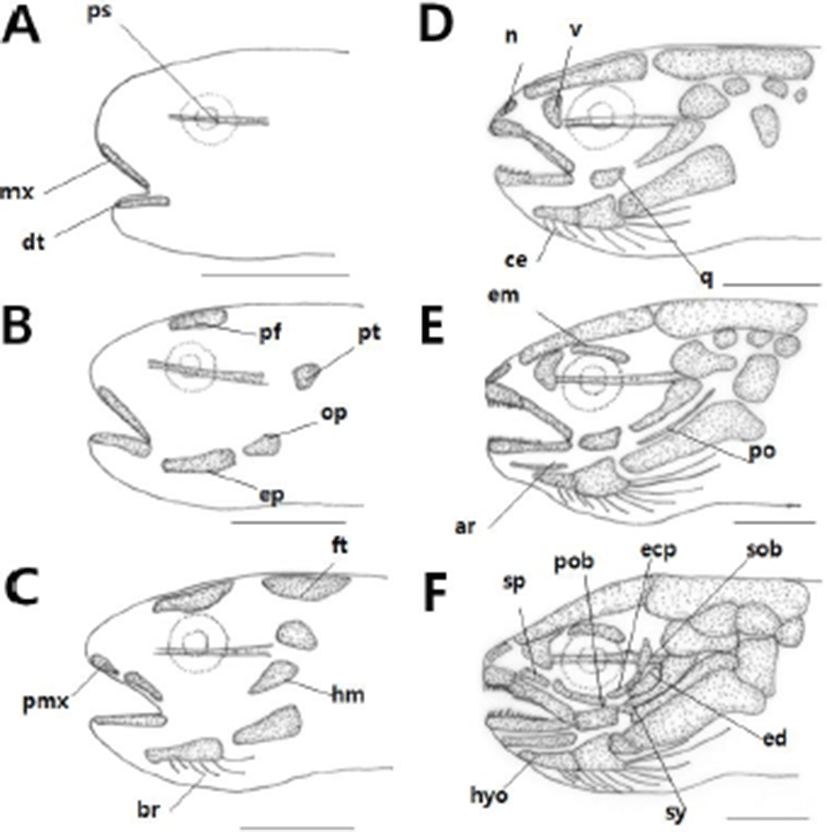
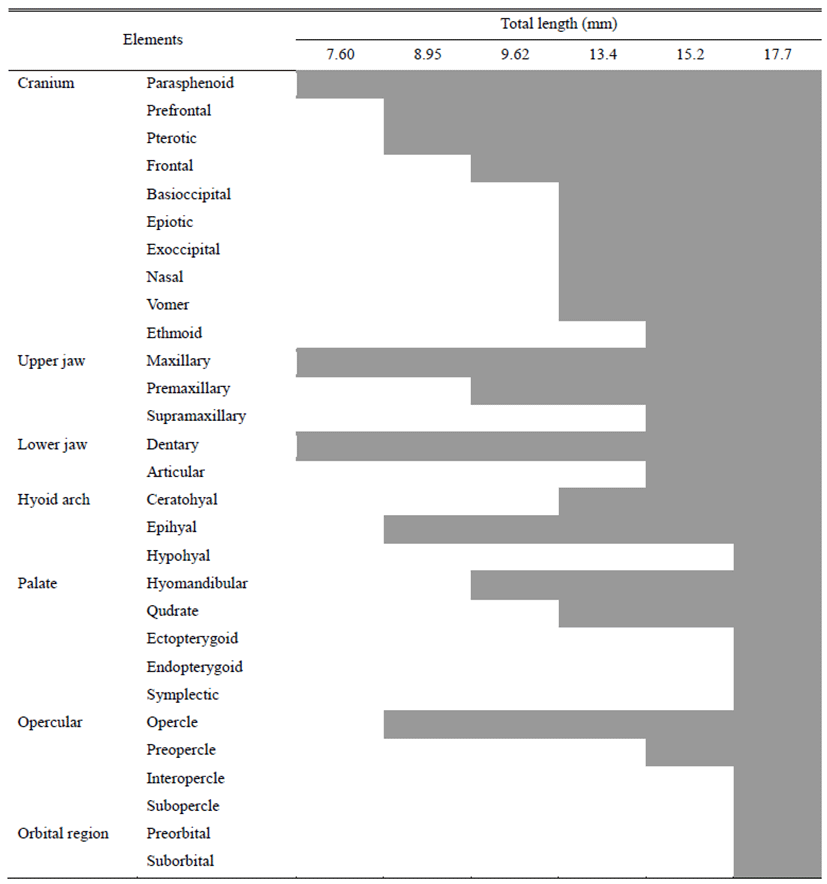
On the 3 day after hatching, Pre-larvae had an average total length of 8.92 to 8.99 (mean 8.95±0.04) mm, and prefrontal ossification in the cranium. Pterotic ossification was observed behind the parasphenoid, epihyal ossified in the hyoid arches of the visceral skeleton, and opercle ossified in the operculum (Fig. 1B).
On the 10th day after hatching, prelarvae were 9.24-10.0 (mean 9.62±0.53) mm in total length and hyomandibular ossified in palate. The frontal ossified in the cranium, and six branchiostegal rays ossified in the hyoid arches. In the jaw bone, the premaxillary was ossified above the maxillary (Fig. 1C).
At 16 days after hatching, posterior larvae were 13.4-13.5 mm in total length (mean 13.4±0.07 mm), and the vomer, nasal, epiotic, exoccipital, posttemporal and basioccipital were ossified in the cranium and the quadrate was ossified in the palate. In hyoid arches, ceratohyal bone was ossified, and branchiostegal rays increased to 9 (Fig. 1D).
At 24 days after hatching, post-larvae were 15.2-15.3 mm in total length (mean 15.2±0.07 mm), and the ethmoid was ossified in the cranium. Supramaxillary and articular ossified in the jaw bone, and preopercle ossified in the operculum (Fig. 1E).
At 30 days after hatching, juvenile was 17.7-17.8 mm in total length (mean 17.8±0.07 mm). The preorbital and three suborbitals were ossified in the cranium orbital region and the endopterygoid and ectopterygoid were ossified. The ossification of cranium and visceral skeleton was all completed with hypohyal ossification in the operculum (Fig. 1F).
Table 2 shows the development of the vertebra, coccyx and pelvic girdle bones of L. obesus larvae. Immediately after hatching, the pre-larvae were 7.60±0.12 mm in mean length and seven centrums in the anterior abdominal vertebrae of the vertebra began to ossify. Five neural spines were simultaneously ossified (Fig. 2A).
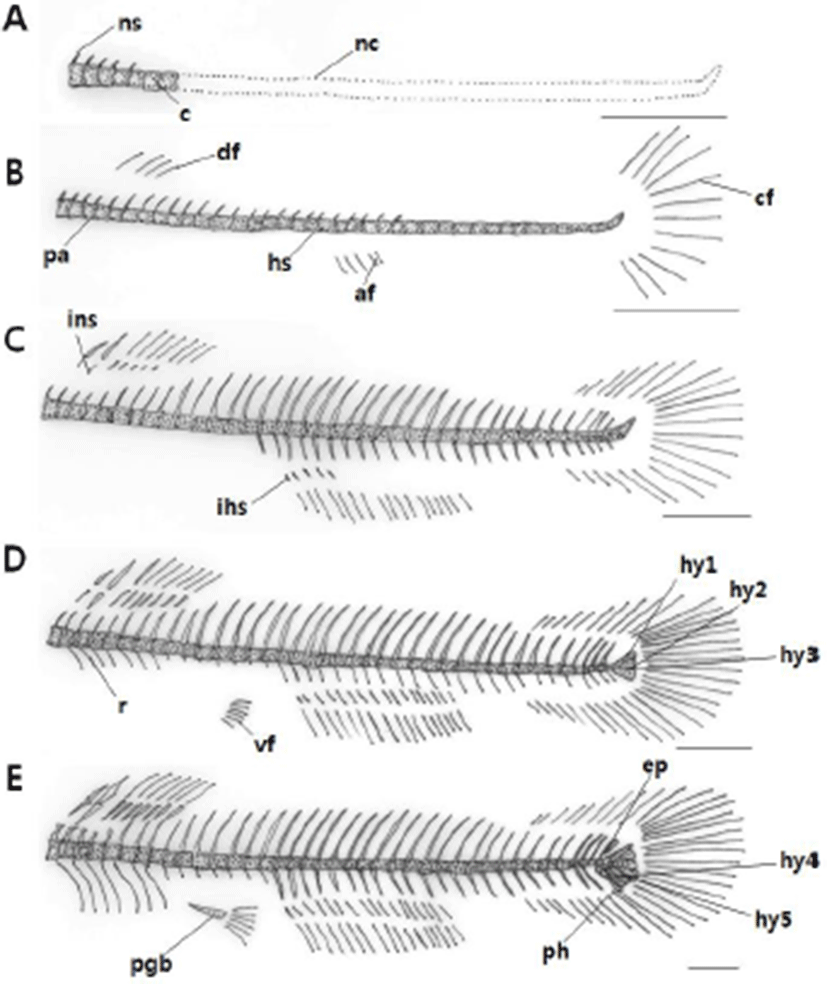
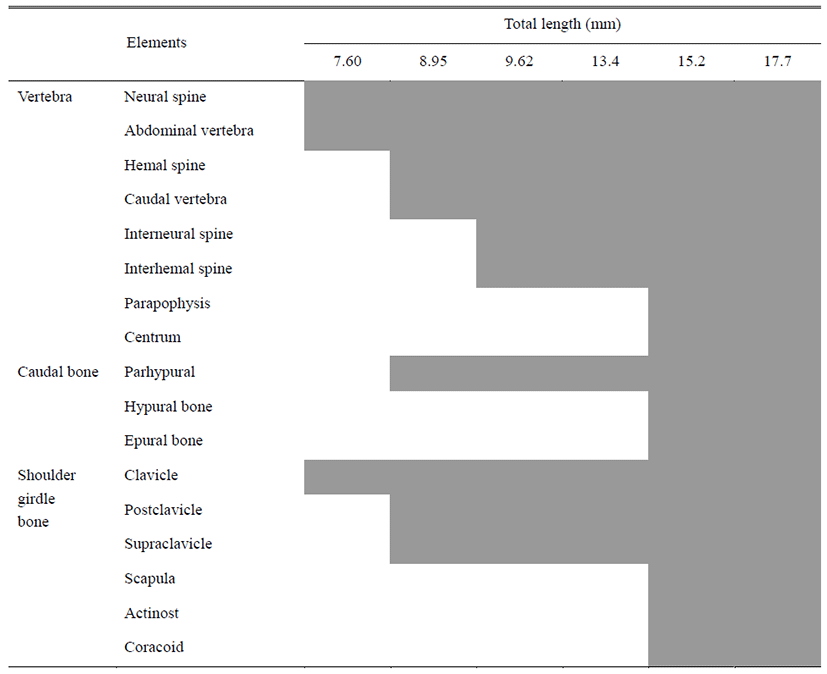
On the 3 day after hatching, the mean length of pre-larvae was 8.95±0.04 mm, and the number of abdominal vertebrae increased to 14. Three parapophysis developed from the front. 24 caudal vertebrae were ossified, and the number of vertebra was 14+24=38. Neural spines were increased to 24, and four hemal spines ossified (Fig. 2B).
On the 10th day after hatching, pre-larvae were 9.62± 0.53 mm in total length and the caudal vertebrae were increased to 28 and the number of vertebra was 14+28=42 to complete ossification. The number of parapophysis was increased to 5, and 7 interneural spines and 5 interhemal spines were ossified (Fig. 2C). On the 16th day after hatching, post-larvae showed an average total length of 13.4± 0.07 mm and an increase of 9 interneural spines. The number of interhemal spines increased to 16-18. During this period, seven ribs began to ossify. Three hypural bones ossified in coccyx (Fig. 2D).
At 24 days after hatching, post-larvae had an average total length of 15.2±0.07 mm, and an epural bone ossified in coccyx. The parhypural bone was ossified, and two pieces of hypural bone were increased to form a total of five fragments. At this time, ossification of coccyx and pelvic girdle bone was completed with the ossification of pelvic girdle bone (Fig. 2E).
In the shoulder girdle bone, the posttemporal is articulated to the cranium and basioccipital, and supports the pectoral fin. The development of shoulder girdle bone of L. obesus larvae is shown in Table 2.
Immediately after hatching, the pre-larvae had an average total length of 7.60±0.12 mm, and the collarbone was ossified first (Fig. 3A). On the 3 day after hatching, the pre-larvae showed an average total length of 8.95±0.04 mm and the upper clavicle ossified at the upper part of the clavicle. And the posterior clavicle was ossified in the lower part (Fig. 3B).
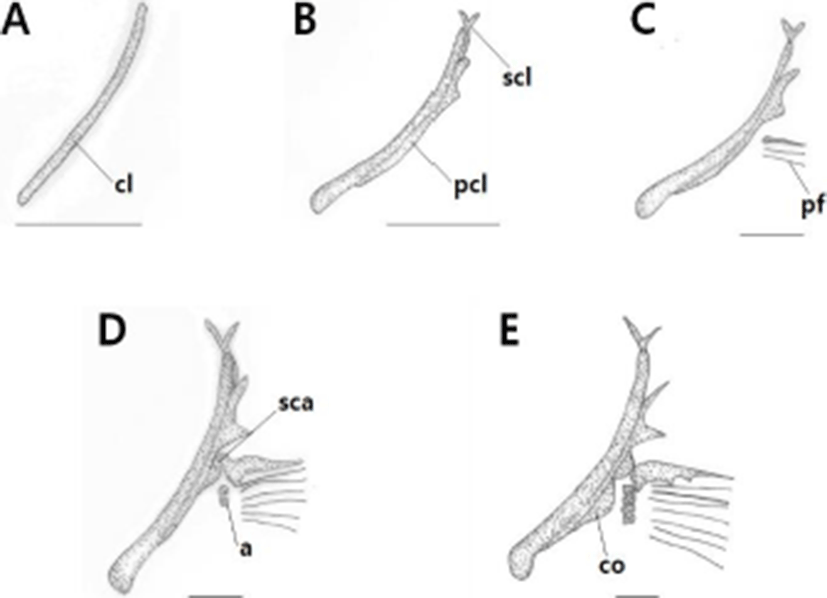
On the 10th day after hatching, pre-larvae had an average total length of 9.62±0.53 mm, and three pectoral stems developed. (Fig. 3C). On the 16th day after hatching, an average total length was 13.4±0.07 mm, and the scapula and two radial bones ossified. The posterior clavicle was elongated (Fig. 3D).
At 24 days after hatching, the post-larvae had an average total length of 15.2±0.07 mm, and coracoids and four radial bones were ossified. The soft ray was increased to 9, and the ossification of the shoulder was completed (Fig. 3E).
DISCUSSION
Interest and precise knowledge of the developmental process of the larvae in the production of fish seeding is essential for the detection and elimination of skeletal abnormalities in the early stages of breeding (Koumoundouros et al., 1997a,b). It provides a basic phylogenetic taxonomic trait of the young stage, and there is a high heat to study this field (Mook, 1977; Potthoff et al., 1988; Faustino & Power, 1999; Liu, 2001; Sfakianakis et al., 2004). In addition, in terms of skeletal development of tibia fish larvae, the jaw bone or visceral skeleton related to food intake and respiratory function have a tendency to be predominantly ossified (Ozawa, 1976). The number of jaw teeth and pharyngeal used for food intake increases with the change in feeding habits (Kohno et al., 1983).
L. obesus was ossified on the first day after hatching, and maxillary and dentary functioning related to cranium parasphenoid and feeding, and clavicle supporting pectoral fin were ossified first. Compared with other fish belonging to other families, Luciogobius guttatus (Kim et al., 1992) showed that the parasphenoid, basioccipital, maxillary, dentary, articular and clavicle were ossified first on the 11th day after hatching. Hyporhamphus sajori (Lee et al., 2001) showed the parasphenoid and clavicle were ossified immediately after hatching. Chaenogobius laevis (Kim & Han, 1989) showed that the basioccipital and parasphenoid of cranium and maxillary of visceral skeleton were ossified at 9-10 days after hatching. The clavicle ossified on the 13th day after hatching.
On the 3 day after hatching, Gasterosteus aculeatus (Han & Kim, 1989) showed that maxillary and dentary including exoccipital, basioccipital, and frontal were ossified. On the 19th day after hatching, the clavicle and upper clavicle were ossified. Rhinchocypris oxycephalus (Han & Noh, 2001) showed that parasphenoid at 6 days after hatching and the clavicle ossified at 13 days after hatching.
While Lampris guttatus (Kim et al., 1992) and H. sajori (Lee et al., 2001) as well as L. obesus showed simultaneous ossification of parasphenoid, jaw bone, and clavicle, the Chysis laevis (Kim & Han, 1989), Gasterosteus aculeatus (Han & Kim, 1989) and R. oxycephalus (Han & Noh, 2001) showed that parasphenoid and jaw bone first began to ossify followed by the clavicle. This showed a difference in skeletal development pattern. However, in all of these species, the jaw bone constituting the jaw is first ossified, suggesting that the cranium is an important element for feeding and respiration, and is the site where ossification occurs preferentially (Vandewalle et al., 1997). At the same time, the clavicle supporting the sternohyoideus muscle plays an important role in the mouth opening and function of feeding (Wagemans & Vandewalle, 1999).
According to the shape development at the larvae stage, the development of the internal skeleton, especially the formation of thorns at the craniofacial skeleton, varies depending on the developmental condition and environment, but thorns formed in the parietal region, operculum, orbit, and shoulder girdle are important traits (Okiyama, 1988). L. obesus has one spinous ray in the pectoral fin of the shoulder girdle, very sharp and hard, with three to five small thorns. The same Amblycipitidae fish, L. mediadiposalis (Choi et al., 2008), has four to six thorns on the pectoral spinous ray. L. andersoni has one to three thorns, which is an important trait to distinguish these three species. Meanwhile, the shape of the body is similar to each other at the young fish stage, and the number and timing of thorns formed on the spinous ray of pectoral fin differs from species to species, which can lead to confusion in classification. On the 24th day after hatching, L. obesus showed that two thorns of the spinous ray of pectoral fin were formed for the first time at an average total length of 15.3 mm. L. mediadiposalis showed that two to three thorns were formed at an average total length of 16.3 mm on the 35th day after hatching, and the number of thorns is similar, but L. mediadiposalis (Choi et al., 2008) was found to have one more than L. obesus In addition, the formation timing for L. mediadiposalis (Choi et al., 2008) was later than L. obesus.
Among the skeletons that support the pectoral fins, a rather large foramen is formed in the scapula, which is considered to be a typical trait that occurs mainly in sea anemone fish (Koumoundouros et al., 2001). On the 24th day after hatching to the juvenile, and when the average length was 15.3 mm, the ossification of the shoulder girdle of L. obesus is complete, and no holes were formed in the scapula in this period. Spotted sea bass, Lateolabrax maculatus (Kang et al., 2012) showed that holes were formed in the scapula and osteochondral bone, but the holes formed in the osteochondral bone especially disappeared as it grew. Based on the fact that these are formed in many Scorpaeniformes fishes (Kim et al., 1997; Han et al., 2001), including many perciformes fishes such as Dentex tumifrons (Koumoundouros et al., 2001), and damsel fishes (Emery, 1973), it is likely that it is a common trait of fish. L. obesus differs from other fish in that there is no hole formed in the scapula. To confirm whether or not the typical traits in Amblycipitidae fish are judged, it is necessary to carry out research on the development of larvae skeleton of L. mediadiposalis and L. andersoni.
In terms of the vertebra ossification of L. obesus larvae, seven centrums were ossified in abdominal vertebrae with an average total length of 7.92 mm. The ossification progressed from the abdominal vertebrae to the caudal vertebrae. After that, at the same time as the ossification of the centrum was completed, the ossification of the tibia was completed. C. laevis (Kim & Han, 1989) showed that the caudal peduncle was ossified after the ossification of the centrum was complete. As for L. guttatus (Kim et al., 1992), H. sajori (Lee et al., 2001), R. oxycephalus (Han & Noh, 2001), Pungtungia herzi (Lee et al., 2002), Pseudopungtungia nigra (Lee, 2004) before the ossification of the centrum was completed, the caudal peduncle was ossified, showing a difference.
Thus, in the skeletal formation of fish, ossification is controlled by their lifestyle, and the difference in lifestyle leads to changes in the degree and order of ossification, which can make various changes in skeletal development of larvae (Mook, 1977). In addition, studies on skeleton can be used as useful data in ecology as well as species identification. It is also said that it is useful to understand the skeletal structure of fishes and to get systematic estimation, and so therefore, it is necessary to continue research on skeletal development in the future.