
INTRODUCTION
Luteinizing hormone (LH) is secreted by the anterior pituitary and plays a role in reproductive functions such as ovulation in females and synthesis of androgens in males (Ezcurra & Humaidan, 2014). Furthermore, LH is a versatile molecule that might play a various role and has been reported to be expressed in extrapituitary regions (Harvey et al., 2012). For example, LH is found in the fetal brain and persists in the adult rat and human (Hojvat et al., 1982; Bowen et al., 2004), in rat testis (Zhang et al., 1995a; Kim & Lee, 2017), in hamster testis (Choi, 2022), in ovine pineal gland (Noteborn et al., 1992), and in cycling rat uterus (Kim & Lee, 1999).
Concerning the placenta, which exists temporarily during pregnancy and connects the mother and fetus, it is well-known that placenta secretes a large amount of chorionic gonadotropin (CG) which has a structure very similar to LH in some mammalian species including rat (Blank & Dufau, 1983; Herkert et al., 2022). In this regard, it is interesting that, even though CG exists as a substitute for LH, transcripts identical to the LH subunits found in the pituitary gland are also present in rat placenta (Shinozaki et al., 1997).
Research on the placental LH and/or LH-like substance is potentially important as it is a physiological signaling molecule with great potential to play a crucial role in rapid tissue development and differentiation after implantation. As on the first try, we performed immunohistochemistry (IHC) to investigate the tissue distribution of the mice placental LH like substances, a common alpha subunit (Cgα) of glycoprotein hormones, LH beta subunit (LH-β) and LH receptor (LH-R).
MATERIALS AND METHODS
All animal experiments were approved by the Institutional Animal Care and Use Committees (IACUC; approval number: E-1302-3). Timely pregnant mice on gestation day (GD) 12 were supplied by the DBL (Eumseong, Korea) and raised for 5 days at the Sangmyung University animal facility under a 12 hours light/dark cycle (lights on at 07:00 hour), with food and water supplied ad libitum at constant temperature of 22±1°C. The animal was sacrificed on GD 17.
After sacrificing the mouse, the placental-fetal pairs were rapidly removed. For tissue section preparation, placentae were immediately immersed in 4% paraformaldehyde (PFA, Bio PURE, Hampshire, UK) in phosphate buffered saline (PBS, pH 7.4, Bio PURE) and stored at 4°C for 24 hours.
Fixed placentae in 4% PFA were dehydrated in graded ethanol (70%, 80%, 90%, 95%, 100%; Duksan, Ansan, Korea) each for an hour and a half using gentle agitation (Green SSeriker, Gwangju, Gyeonggi-do, Korea) and then in absolute ethanol (Merck, Darmstadt, Germany) overnight. After that, the samples were immersed in xylene (Samchun, Pyeongtaek, Korea) at room temperature 3 times at intervals of 40 minutes and into paraffin (Merck) at 60°C 3 times at intervals of 20 minutes. Tissue was embedded (Microm, Walldorf, Germany) in paraffin using the cassette (Hyunil LAB-MATE, Seoul, Korea). The block was sectioned using microtome (Microm) to a thickness of 6 μm and specimens were attached to microscope slides (Marienfeld, Lauda-Königshofen, Germany) using water bath (Microm). Heat was applied to the specimens at 63°C in the slide warmer (Jisico, Seoul, Korea) for 12 hours.
The slides were dewaxed in xylene for 5 minutes, 3 times and hydrated in 100%, 95%, 90%, 80%, and 70% ethanol for 5 minutes each. The specimens were washed in PBS for 5 minutes, 4 times and immersed in hydrogen peroxide solution [10% H2O2 (Daejung, Siheung, Korea) and 10% Methanol (Merck)] in PBS for 20 minutes, and washed in PBS 2 times. The slides were incubated with blocking serum [2% BSA (Bio PURE) and 2% normal goat serum (VECTOR, Newark, CA, USA)] in PBS for 1 hour and incubated overnight with diluted primary antibody (dilution of 1:50, 1:500 and 1:1,000, respectively, for Cgα, LH-β, and LH-R). Antisera for Cgα and LH-β were purchased from National Hormone & Pituitary Program (NHPP, Torrance, CA, USA) and antiserum for LH-R was purchased from Mybiosource (MBS244081, San Diego, CA, USA). The slides were washed 2 times and incubated with biotinylated goat anti-rabbit IgG secondary antibody (VECTOR) for 1 hour (in the case of Cgα, overnight) and washed in PBS 2 times. The samples were incubated with avidin-biotin-peroxidase complex using the ABC kit (VECTOR) for 40 minutes and washed 2 times. The chromogenic reaction was progressed using the DAB kit (VECTOR) and stopped by washing in tap water. The slides were stained in hematoxylin (VECTOR) for 7 minutes and dehydrated in graded ethanol (70%, 80%, 90%, 95%, and 100%) for 3 minutes each and then cleaned in xylene for 5 minutes, 3 times. The slides were mounted with Permount (Fisher Scientific, Waltham, MA, USA). The images were captured with the BX51 Microscope & DP70 Digital Camera System (Olympus, Tokyo, Japan).
RESULTS
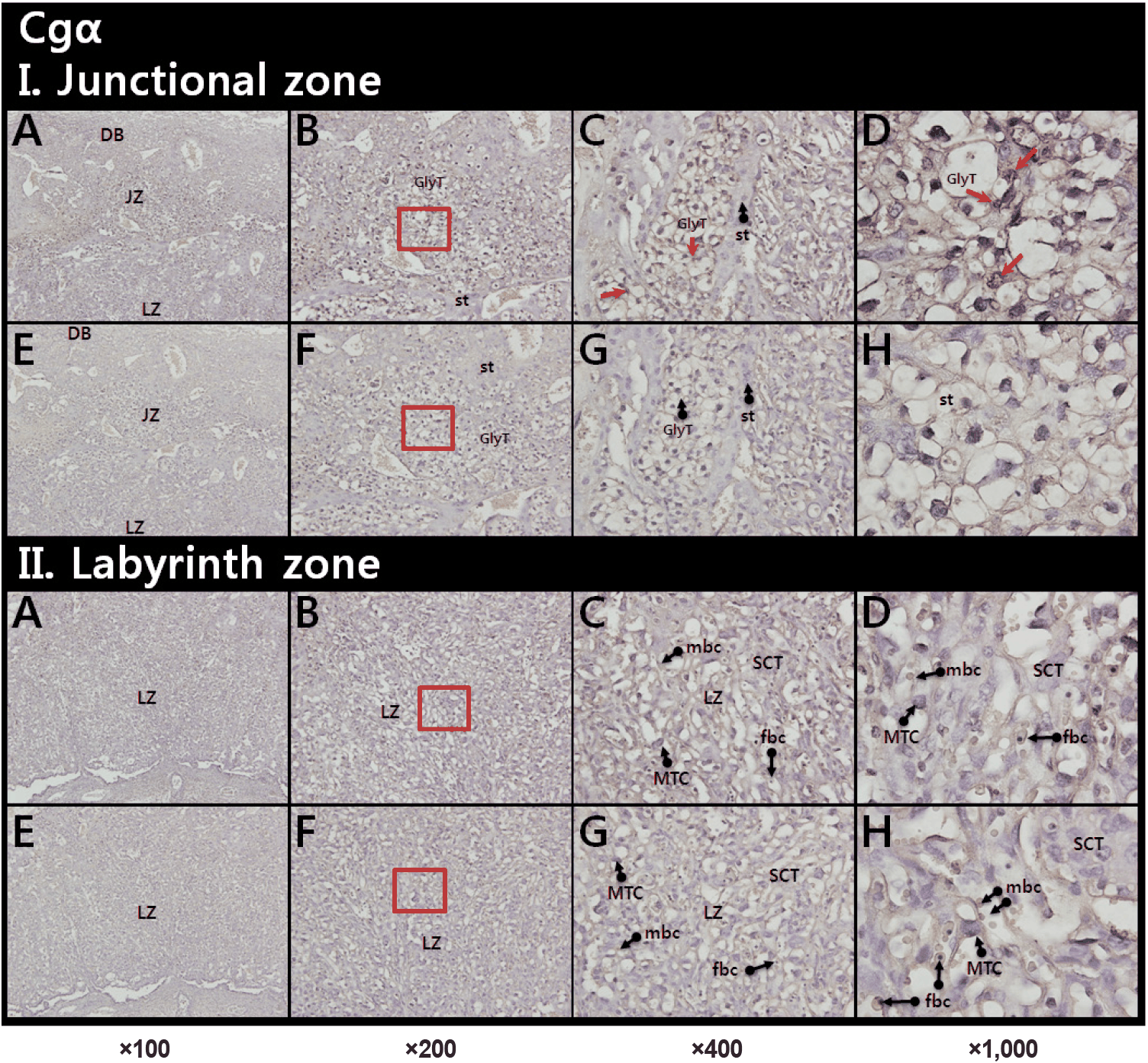
Localization of Cgα, LH-β, and LH-R in the mouse placenta was investigated by specific IHCs. Cgα staining were detected on the nuclei of glycogen rich trophoblast cells (GlyT, red arrows) in a junctional zone (JZ; Fig. 1, upper panel). In a labyrinth zone (LZ), weaker signals were found in mononucleate trophoblast cell, and nuclei of fetal blood cell (fbc) (Fig. 1, lower panel).
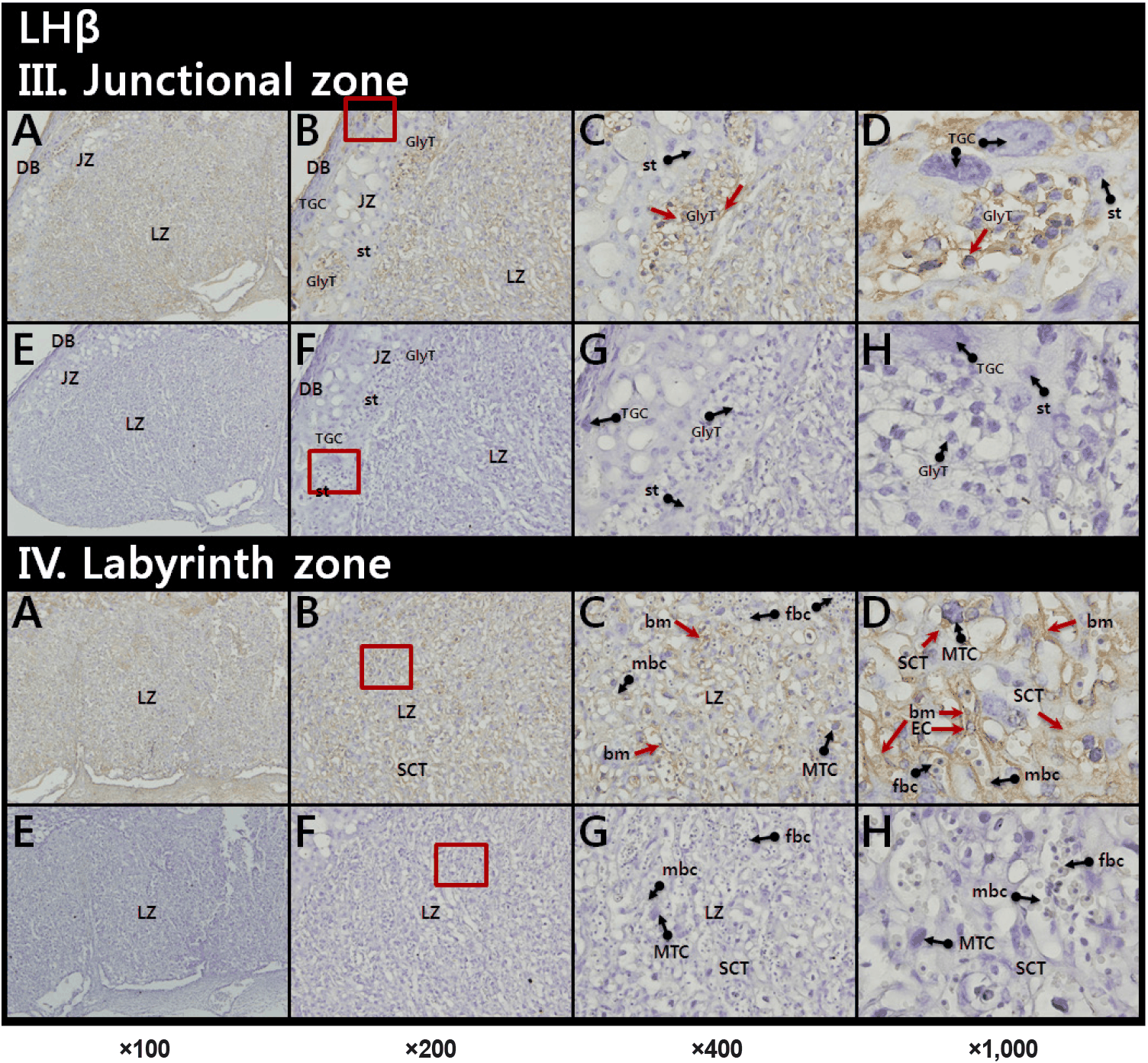
LH-β staining was observed in all cell layers. Especially, immunoreactive LH-β was detected in and around GlyT in the JZ (Fig. 2, upper panel) and on cytoplasm of syncytiotrophoblast cells (SCT), fetal capillary endothelial cells (EC), and basement membrane (bm) in the LZ (Fig. 2, lower panel).
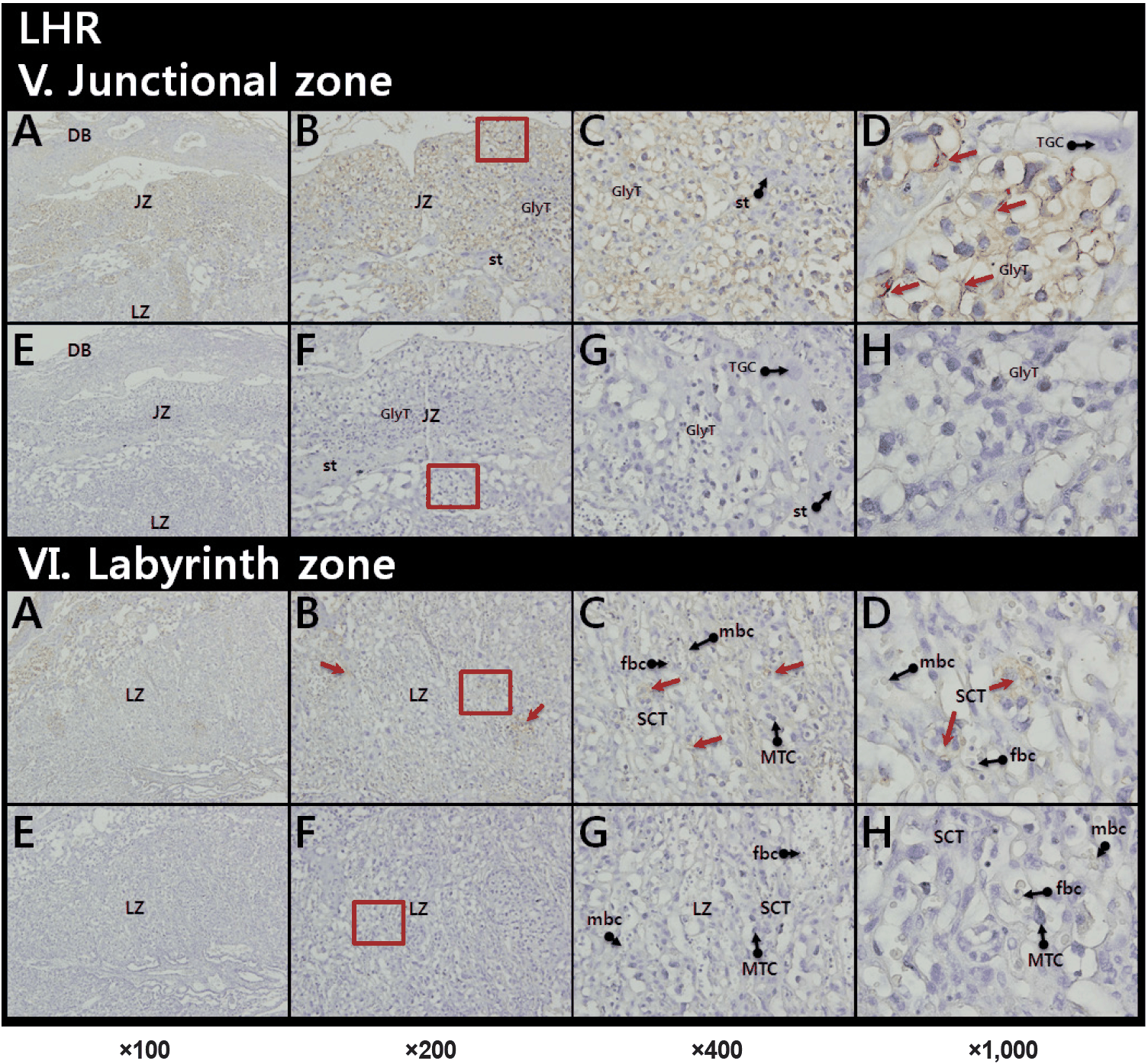
Our IHC also revealed that the immunoreactive LH-R located on the membrane and in a part of cytoplasm of GlyT in the JZ (Fig. 3, upper panel). Relatively weak LH-R staining was also shown in the SCT layer of the LZ (Fig. 3, lower panel).
DISCUSSION
Research on LH expression outside the pituitary gland has been ongoing for decades, but little amount of information is available so far. Extrapituitary production of LH has been reported primarily in reproductive organs like gonads, as well as the accessory sex organs (Harvey et al., 2012); The local LH has been identified in the rat testis (Zhang et al., 1995a; Kim & Lee, 2017), mouse testis (Kim & Lee, 2017), hamster testis (Choi, 2022), rat ovary (Schirman-Hildesheim et al., 2008), rat uterus and epididymis (Kim & Lee, 1999; Lee & Lee, 1999), and rat placenta (Shinozaki et al., 1997). This local production, particularly in gonads, might acts in a paracrine or autocrine manner to modulate gametogenesis, steroidogenesis and other reproductive-related phenomena. For example, while the pituitary provides the main endocrine surge for ovulation, the local LH might act as a ‘fine-tuning’ mediator of reproductive signals.
The placenta is a temporary organ that forms inside the uterus during pregnancy to connect the mother and the fetus. It performs respiration, nutrient supply, and excretion functions essential for the survival and growth of the fetus, and is expelled from the body after childbirth (Wide & Wide, 1979; Makrigiannakis et al., 2017; Herkert et al., 2022). HCG is a hormone secreted by the placenta in early pregnancy that maintains corpus luteum function and helps produce progesterone to maintain pregnancy (Cindrova-Davies & Sferruzzi-Perri, 2022). Interestingly, HCG has structure and function which are very similar to LH, therefore, one could wonder why pituitary type LH subunits are expressed in the placenta (Shinozaki et al., 1997).
The gonadotropin (LH, FSH, and hCG) is noncovalently joined by the common α subunit and specific β subunit (Ryan et al., 1988; Alevizaki & Huhtaniemi, 2002). The Cgα gene is coded by a single gene that consists of 4 exons and 3 introns (Gharib et al., 1990). The authentic LH-β gene is composed of 3 exons and 2 introns in the mouse (Kumar & Matzuk, 1995). LH is well known to be synthesized and secreted by the anterior pituitary (Steelman et al., 1959; Hartree, 1966; Lynch et al., 1988). In contrast, Zhang et al. found the expression of testis-specific mRNA variants of the LH-β gene (Zhang et al., 1995a). There are three testicular transcripts in the rat of lengths of 3.2, 2.4, and 0.86 kb (Zhang et al., 1995b). Follow-up studies, including our own, indicates that the presence of tissue-specific expression of LH-β gene in mouse testis (Kim & Lee, 2017), hamster testis (Choi, 2022), and rat ovary (Schirman-Hildesheim et al., 2008).
The presence of transcripts for placental LH subunits in rat was initially proven by reverse transcription-polymerase chain reactions (RT-PCR), Southern blot assay and partial sequencing of the PCR products (Shinozaki et al., 1997). We recently found that the transcripts for mice placental LH-β were almost identical to those found in rat testis including all known exons of authentic pituitary type transcripts (data not shown). So far, the physiological significance of the placental LH subunit substances has not been addressed. Regarding the gonadal and placental LH, research on their molecular structure(s) and characteristics must be conducted first. In particular, it is necessary to investigate molecular properties, such as the molecular weight of partially purified tissue extracts, by adopting classical methodologies such as column chromatography, and measure physiological effects through in vivo injection or in vitro treatment.
The present study demonstrated that immunoreactive LH subunits like substances were localized differentially in the microstructures of mice placentae. The fact that substances exhibiting immune responses exist in the same cells or adjacent cells suggests the possibility of local regulation between these substances. Further studies will be helpful for understanding the biochemical nature and the role(s) of local LH-like substances in sophisticated placental physiology.