
INTRODUCTION
The grass puffer, Takifugu niphobles, is the most commonly found fish in adjacent seas of Korea. Grass puffer inhabits in coastal and brackish waters where sea water and fresh water are mixed with low salinity. They usually eat small crustaceans, shellfish, and cobwebds (Honma et al., 1980; Oh et al., 2000). Grass puffer exhibits unique spawning behaviors. It spawns in semilunar cycles from late spring (May) to summer (August). It will spawn to the intertidal zone and lay eggs in rock slits (Honma et al., 1980; Oh et al., 2000; Masaomi et al., 2013; Goo et al., 2015; Gil et al., 2017b). The grass puffer can grow up to 20 cm. It is the smallest adult fish among Tetraodontidae fish (Nozaki et al., 1976; Oh et al., 2000). Because of its large population and small size, it is widely used as an ornamental fish. It is also possible to replace the expensive puffer fish dishes.
For grass puffer, anesthetic and physiological effects of clove oil and lidocaine-HCl, effect of starvation, sexual dimorphism were previously studied (Gil et al., 2017a; Kim et al., 2017; Park, 2017). However, unlike previous studies, the objective of this study was to determine cleavage frequency and mitotic intervals of cell divisions in grass puffer at different water temperatures. Effective controls of the first cleavage and second ootid for chromosome manipulation in fish are dependent on the type and duration time of treatment. It is well known that the second meiosis is affected by water temperature, hydraulic pressure, and chemical treatment (Thorgaard et al., 1981; Thorgaard & Allen, 1987; Ahn, 1995). Therefore, among different methods of fish breeding techniques, producing polyploidy, gynogenetic, and androgenetic diploid fish through chromosome manipulation is very important for the aquaculture industry (Thorgaard, 1983; Thorgaard & Allen, 1987; Mims et al., 1997). Recently, Masaomi et al. (2013) have induced triploid grass puffer. Yoo et al. (2018) have also produced hybrids and hybrid triploids between river puffer and tiger puffer.
Among different techniques for chromosome manipulation, inhibiting the first cleavage and first or second polar body emission has been adapted to induce tetraploid or mitotic gynogenetic and androgenetic diploid (Mims et al., 1997; Masaomi et al., 2013). An appropriate Dettlaff unit is determined by the duration of one mitotic cycle during the time of synchronous cleavage divisions under different water temperatures. The Dettlaff unit is the duration in minutes of one mitotic cycle during early simultaneously embryonic beginning or the interval between two continuous cell divisions separation (Shelton, 1993; Saat & Veersalu, 1996; Shelton et al., 1997). When measured over a range of temperatures, the relationship between mitotic intervals (τ0) and temperature as determined by regression analysis can be used to predict developmental events that are influenced by temperature within a single species or between species with similar spawning biology (Dettlaff, 1986). Previous studies have been limited to egg development of grass puffer (Oh et al., 2000). Grass puffer is a very useful sample for fishery laboratory animals in physiological, hematological, and other experiments. Therefore, the objective of this study is to determine the mitotic intervals (τ0) of two consecutive cell divisions and synchronous embryonic cleavage in grass puffer at different water temperatures (18, 20, 22, and 24℃).
MATERIALS AND METHODS
Grass puffer, Takifugu niphobles, were collected from the harbor at Korea Maritime and Ocean University (KMOU), Busan, Korea, from May to August, 2016. Samples were transported to and reared in Fishery Genetics and Breeding Sciences Laboratory of the KMOU, Busan, Korea. Initial measurements of body weight (BW) and total length (TL) were made to the nearest 0.1 g and 0.1 cm using an electric balance (AX 200, Shimadzu Corp., Japan) and digital vernier calipers (CD-20 CP; Mitutoyo, Japan), respectively. The TL and BW of each species were 13.6±2.15 cm and 29.7±4.26 g in females and 12.7±2.53 cm and 26.5±3.29 g in males, respectively. Experimental samples were acclimated for one week in aquariums (200 L) with different water temperatures (18, 20, 22, and 24℃) to have sufficient adaptation.
We injected into the grass puffer 1,000 IU/g BW human chorionic gonadotropin (HCG, Sigma-Aldrich St. Louis, MO, USA). Twenty-four hours after the injection, eggs and sperm were extracted by pressing the abdomens. They were mixed in sufficient seawater by drying method. Fertilized eggs were adequately aerated and filtered in plastic sea water tank (20 L). Comparing the size of unfertilized eggs and fertilized eggs, unfertilized eggs were measured at 20℃ while the fertilized eggs were divided into four different groups based on water temperature (18, 20, 22, and 24℃). Color, type, weight, and size (major axis, minor axis, and extracellular matrix) of unfertilized and fertilized eggs (when blastodisc stage was developed) of grass puffers were compared. The weights of eggs were measured by weighing 10 eggs and dividing the number by 10. The diameters of eggs were measured using the eyepiece micrometer of a microscope (Axioskop, Zeiss, Germany) under 50 × magnification. Egg development was confirmed by sampling at intervals of 5 mins from 1 hr after fertilization.
To investigate the period of the first cleavage of fertilized eggs depending on different temperatures also known as the Dettlaff unit (mitotic interval, τ0) of different cleavage frequencies and water temperatures, eggs were fertilized at 18, 20, 22, and 24℃ and sampled every 5 mins after fertilization. Each sample was fixed in 10% neutral formalin (100 mL formalin, 6.5 g Na2HPO4·12H2O, 4 g KH2PO4, fresh water 900 mL) and kept at 4℃ until observation. τ0 was examined under microscope with 50 × magnification. For τ0, the time it took for about 10% of the entire eggs to reach 2-cell stage (τI) and 8-cell stage (τIII) at each hatching temperature were measured according to the method of Ignat’eva (1975). Average τ0 was calculated by the formula τ0=(τIII–τI)/2. Linear regression analysis was performed to find the relationship between the average τ0 and water temperature. This experiment was performed in triplicates.
RESULTS AND DISCUSSION
As shown in Table 1 and Fig. 1, egg color of grass puffer, Takifugu niphobles, is light yellowish. Egg type is unadhesive and demersal. Egg weight was measured at 0.09± 0.002 mg at water temperature of 20℃. Among various puffer fish species, the eggs of grass puffer are light yellowish, unadhesive, and demersal with lots of oil globules (Oh et al., 2000). Regarding other puffer fish species, the eggs of tiger puffer (T. rubripes), river puffer (T. obscurus), and panther puffer (T. pardalis) are opaque and milky-white in color (Jang et al., 1996; Han, 1999; Han et al., 2001). The major axis of unfertilized egg was 1.05±0.041 mm. The minor axis of unfertilized egg was 1.01±0.038 mm. The extracellular matrix of unfertilized egg was 0.09±0.005 mm. The sizes of fertilized eggs at four different water temperatures were significantly smaller than that of the other three groups in the long axis and short axis at 18℃ and no significant difference was found in the other three groups (p<0.05). The sizes of extracellular matrix showed no significant (p>0.05) difference among four water temperature groups. When fertilized eggs were compared to unfertilized eggs at the same temperature of 20℃, the major axis of fertilized egg was 1.36 times larger and the minor axis was 1.39 times larger (p<0.05) than those of unfertilized eggs.
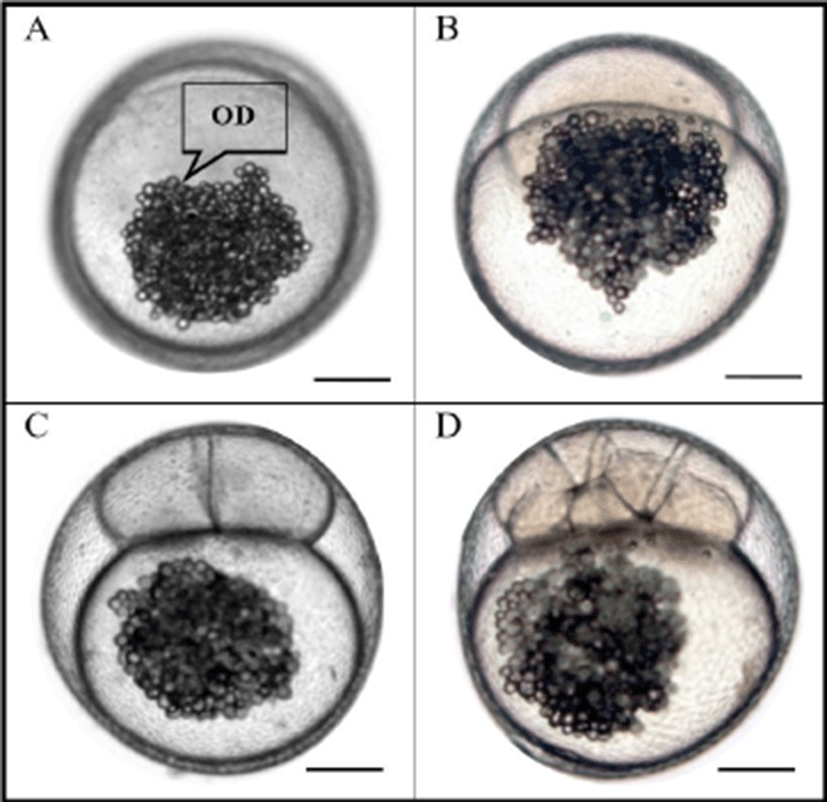
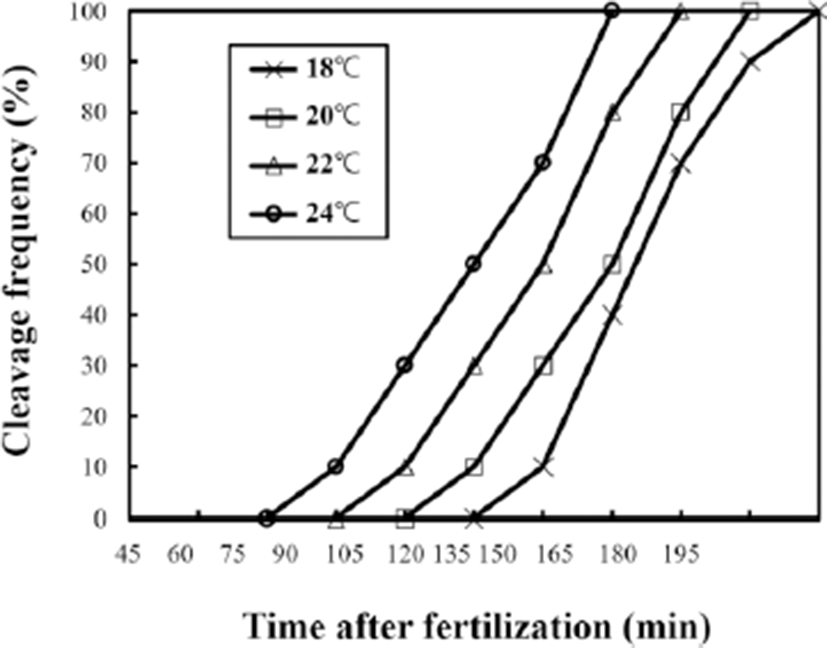
As shown in Fig. 2, egg development was accelerated when water temperature was increased from 18℃ to 24℃. Egg developmental rates of the four water temperature groups were similar to each other. However, the time that eggs reached the first cleavage was 30 mins at 18℃, 45 mins at 20℃, 60 mins at 22℃, and 75 mins at 24℃. After 90 mins, the first cleavage was finished in all water temperature groups. Oh et al. (2000) have reported the embryogenesis and early ontogenesis of a grass puffer. As mentioned by Oh et al. (2000), two-cell stage of grass puffer egg can be reached within two hours after fertilization at 22℃ and 8-cell stage of egg can be reached at 4 hours after fertilization. Although our experimental water temperatures were different from those of Oh et al. (2000), the time needed to reach the 2-cell stage and the 8-cell stage at 18, 20, 22, and 24℃ was similar to that of Oh et al. (2000) at 20℃. Egg development stage of tiger puffer is different from that of grass puffer. For example, the time needed to reach 2-cell stage and 8-cell stage has been reported to be 3:30 and 5:50 hours after fertilization, respectively (Han, 1999). Therefore, there is a time difference in egg development stage among different fish species or the same fish species at different temperatures. This could be used to identify the taxonomic ranges of different fish species. The eggs of grass puffer showed faster development at higher temperature (Fig. 2). As the water temperature was increased, the slope of the first cleavage frequency with elapsed time after fertilization was increased. Approximately 10% of fertilized eggs reached first cleavage frequency every 15 min. These results are similar to those reported by Ignat’eva (1975), Saat & Veersalu (1996), Park & Im (2001), and Kim et al. (2011) showing that faster development has occurred at higher water temperature.
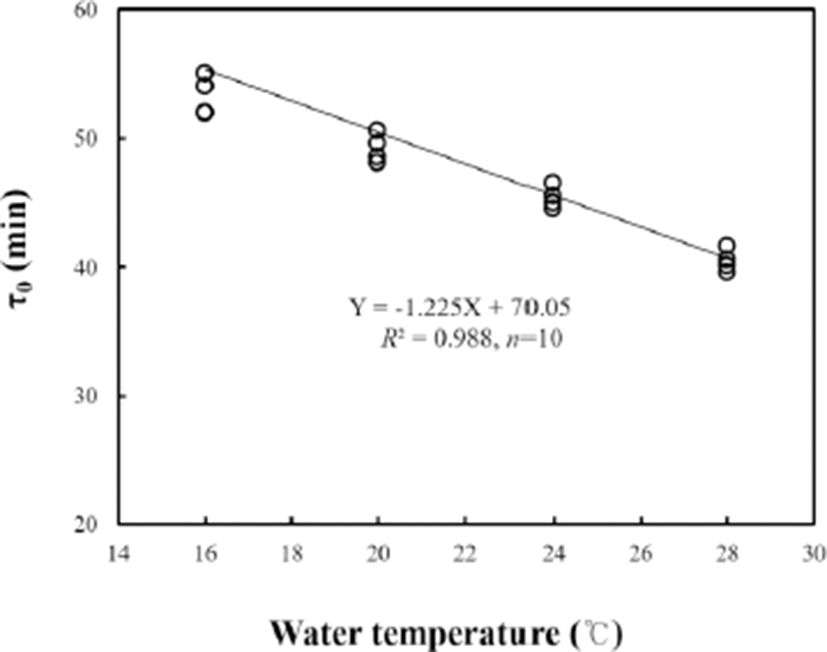
As shown in Fig. 3, the time of mitotic intervals (τ0) was measured according to water temperature (18, 20, 22, and 24℃). Unlike the tendency of cleavage frequency, τ0 was negatively correlated with water temperature (Y=–1.225X+ 70.05, R2=0.988 where Y was τ0 and X was temperature). Of the four water temperature groups, τ0 was 53±1.9 mins at 18℃, 49±2.3 mins at 20℃, 45±1.7 mins at 22℃, and 40±1.8 mins at 24℃. The duration of one mitotic cycle during the period of identical cell divisions (τ0) has proven to be an appropriate unit for comparing the duration of development processes at different temperatures in poikilothermic animals undergoing identical cell divisions during their early development (Shelton, 1993, 1997; Park & Im, 2001; Kim et al., 2011). According to results shown in Fig. 3, we found that grass puffer showed faster development with decreased mitotic intervals when water temperature was increased, indicating a strong negative correlation between τ0 and water temperature. Besides this study, similar results on the reduction of mitotic interval according to rising water temperature have been reported in far eastern catfish (Silurus asotus), black plaice (Pleuronectes obscurus), and Korean rose bitterling, Rhodeus uyekii (Park & Im, 2001; Park and Im, 2010; Kim et al., 2011).
In summary, this study demonstrated obvious specific and clear differences in the time of the first cleavage and mitotic intervals at different temperatures in grass puffer. Data obtained from this study will add value to biological Takifugu species aquaculture database. This study will aid future investigations on seeding production by artificial fertilization, induction of triploid for producing sterile organisms, and induction of tetraploid, mitotic gynogenetic diploid, and androgenetic diploid grass puffer.