INTRODUCTION
Cynoglossus spp. is, biologically warm water teleost species, belonging to family Cynoglossidae, order Pleuronectiformes, class Actinopterygii, widely distributed on the coast of the Yellow Sea, southern sea in the Korean Peninsula, as well as in several areas in East China and the Bo Hai of China. Under the natural ecosystem, the tonguesoles inhabit in the flats consisting of a lot of mud, silt and slime in the coast where the clams, crabs and the other shellfishes are abundant. Tonguesole is one of the most popular marine products and economically important aquacultural species in Korea because of their taste and nourishing value, and Koreans consume them plentifully. The tonguesoles live buried in sandy mud at a number of depths. However, in spite of their economic and scientific consequences, a little information currently exists regarding the physiological and ecological levels of tonguesole species in Korea. Nevertheless, this tonguesole, which is distinguished important reproductively (Lee et al., 2000; Kang et al., 2012; Lim et al., 2016), morphologically (Choi, 1996; Gu et al., 2009; Kwun & Kim, 2010), as well as ecologically (Choi et al., 1995; Baeck & Huh, 2004) are a few genetically and/or molecular-biologically (Kwun & Kim, 2010; Noh et al., 2016) studied like other finfishes. There are more wishes to understand the genetic traits and conformation of this fish population in order to evaluate precisely the genetic consequences.
The specific markers particular to species, genus or geographical population have all been employed in the identification and discrimination of individuals, species and populations, hybrid parentages and genetic diagnostics (Callejas & Ochando, 1998; Huang et al., 2000; Kim et al., 2000; Yoon & Kim, 2004). Thus, the applications of PCR techniques to aquaculture had been to identify genetic distances among a few of fish species and/or invertebrates apart from geographic sites (Partis & Wells, 1996; Callejas & Ochando, 1998; McCormack et al., 2000; Kwun & Kim, 2010; Song & Yoon, 2013; Oh & Yoon, 2014; Yoon, 2015; Noh et al., 2016). In the present study, to elucidate the genetic distances and differences between geographical tonguesole populations, the author performed a clustering analysis of two tonguesole populations collected from two sites of Gunsan and the Atlantic.
MATERIALS AND METHODS
Muscle tissues were collected separately from Gunsan and the Atlantic tonguesole populations, respectively. These tonguesole muscles were collected in sterile tubes, immediately placed on ice, and stored at −80°C until necessary. PCR analysis was performed on the muscle extracts from 22 individuals, using six oligonucleotides primers. DNA extraction should be performed according to the separation and extraction methods as described previously (Song & Yoon, 2013). 600 μL of chloroform was added to the mixture and then inverted (no phenol). After washing several washings, samples of muscle tissues were placed into 10 mL test tubes, to which 3 volumes of lysis buffer (155 mM NH4Cl, 10 mM KHCO3, 1mM EDTA) was added, and the mixture tubes were lightly inverted. The precipitates obtained were diffused with lysis buffer (10 mM Tris-HCl, pH 8.0, 10 mM EDTA, 100 mM NaCl, 0.5% SDS). Samples were added 15 μL proteinase K solutions (10 mg/mL). After incubation, there was added 300 μL of 6 M NaCl and softly pipetted for a few of min. The DNA pellets were incubation-dried for 2 hrs, held at −80°C until analysis, and then dissolved in the TE buffer (10 mM Tris-HCl, pH 8.0; 1 mM EDTA). Ice-cold 70% ethanol was added, and then the samples were centrifuged at 19,621 g for 5 minutes to extract DNA from the lysates. The concentration of the extracted genomic DNA was measured by the optical density at 260 nm, with a spectrophotometer (Beckman Coulter, Buckinghamshire, UK). The DNA pellets were then incubation-dried for more than 12 hours, maintained at −80°C until needed and then dissolved in the distilled water.
The oligonucleotides primers were purchased from Operon Technologies, USA. The OPA-11 (5’-CAATCGCCGT-3’), OPB-09 (5’-TGGGGGACTC-3’), OPB-14 (5’-TCCGCTCTGG-3’), OPB-20 (5’-GGACCCTTAC-3’), OPC-14 (5’-TGCGTGCTTG-3’) and OPC-18 (5’-TGAGTGGGTG-3’) were shown to yield the unique shared loci to each population and the number of shared loci by the two populations which could be clearly scored. The author used these oligonucleotides primers to determine the genetic variations, DNA polymorphisms, and similarity of the tonguesole from Gunsan and the Atlantic. PCR process was performed using a Programmable DNA Thermal Cyclers (MJ Research Inc., Waltham, MA, USA). DNA amplification was performed in 25 μL samples, which contained 10 ng of template DNA, 20 μL of premix (Bioneer Corp., Daejeon, Korea), and 1 unit of primer. Amplification products were generated via electrophoresis on 1.4% agarose (Bioneer Corp., Daejeon, Korea) gel containing TBE (90 mM Tris, pH 8.5; 90 mM borate; 2.5 mM EDTA). The 100 bp ladder marker (Bioneer Corp., Daejeon, Korea) was utilized as a DNA molecular weight marker. The electrophoresed agarose gels were illuminated by ultraviolet rays, and photographed using a photoman direct copy system (PECA Products, Beloit, WI, USA).
Similarity matrix including Bandsharing (BS) values were calculated according to the presence/absence of amplified products at specific positions in the same gel from the DNA profiles. The degree of variability was calculated by use of the Dice coefficient (F), which is given by the formula: F=2 nab / (na+nb), where nab is the number of bands shared between the samples a and b, na is the total number of bands for sample a and nb is the total number of bands for sample b (Jeffreys & Morton, 1987; Yoke-Kqueen & Radu, 2006). The average of within-population similarity was calculated by pairwise comparison between individuals within a population. The relatedness between different individuals in the tonguesole populations was generated according to the bandsharing values and similarity matrix. A hierarchical polar dendrogram was assembled using similarity matrices to yield a cluster tree, which was supported by the Systat version 10 (SPSS Inc., Chicago, IL, USA). The Systat software was also used to analyze genetic distances, Euclidean distances within and between tonguesole populations, means, standard errors, and t-test scores.
RESULTS AND DISCUSSION
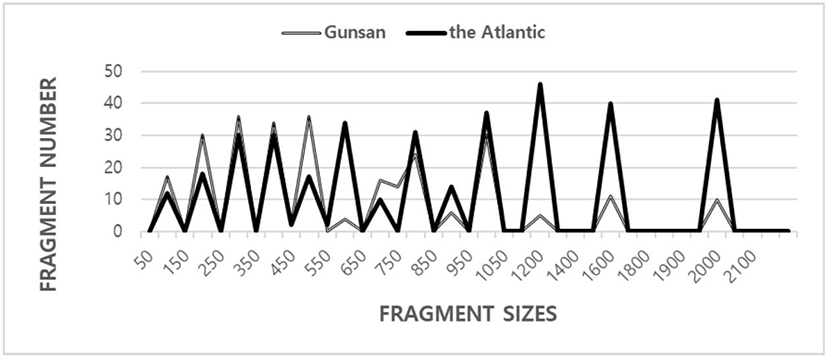
Here, the six oligonucleotides primers OPA-11, OPB-09, OPB-14, OPB-20, OPC-14, and OPC-18 were used to generate the unique shared loci to each population and shared loci by the two populations. The complexity of the banding patterns varied dramatically between the primers from the two zones. The size of the DNA fragments also varied widely, from 100 to 2,050 bp (Fig. 1). The fragment numbers in each size interval have been computed from the integrated fragments attained with every six oligonucleotides primers. The higher fragment sizes (>2,100 bp) are not observed in the two C. spp. populations. Eighty fragments ranging from 200 bp to 2,200 bp were unambiguously counted in the black tiger shrimp (Penaeus monodon) (Tassanakajon et al., 1998). The DNA fragments obtained using the four primers ranged from 100 to 2,300 bp in the brittle star (Amphiura filiformis) (McCormack et al., 2000). The bandsharing value between individuals no. 01 and no. 02 was 0.962, which was the highest value identified within the Gunsan tonguesole population. The bandsharing value between individual’s no. 11 of the Gunsan tonguesole and no. 20 of the Atlantic tonguesole was 0.204, which was the lowest observed. The 22 unique shared loci to each population generated by OPA-11 oligonucleotides primer were in Gunsan tonguesole population, as summarized in Table 2. Excitingly, the oligonucleotides primer OPC-14 generated 44 unique shared loci to each population, which were identifying each species in the Atlantic population (Table 2). Especially, the oligonucleotides primer OPC-18 generated 33 unique shared loci to each population, which were classifying each population in the Atlantic tonguesole. Unusually, the primer OPB-20 detected 22 shared loci by the two populations, which were identical in all samples, as summarized in Table 2. The oligonucleotides primer OPC-18 also detected 66 shared loci by the two populations, major and/or minor fragments of 400 bp, 800 bp, and 1,600 bp in all samples.
| Population | Gunsan | Atlantic |
|---|---|---|
| Gunsan | 0.799±0.016a | 0.452±0.016c |
| Atlantic | - | 0.765±0.011b |
Each value is a result of three different experiments.
a-c Values with different superscript are significantly different, p<0.05.
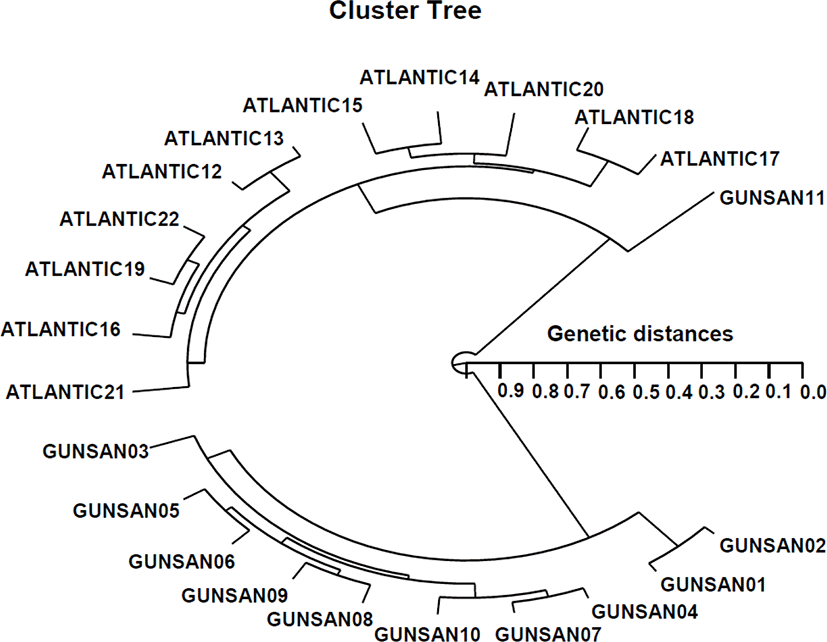
Polymorphisms are determined by the banding patterns of primer-amplified products at specific positions (Tassanakajon et al., 1998; Nozaki et al., 2000; Yoon & Kim, 2001). Several researchers have studied the sizes of DNA fragments in the PCR profiles of five species of Eastern Pacific abalone (genus Haliotis) (Muchmore et al., 1998), black tiger shrimp (Penaeus monodon) (Tassanakajon et al., 1998), the brittle star (Amphiura filiformis) (McCormack et al., 2000) and wild shrimp populations (Yoon & Kim, 2003a). The specific primer proved useful in the identification of individuals and/or populations, resulting from variations in DNA polymorphisms among individuals/populations (Liu et al., 1998; Yoon & Kim, 2003b; Kim et al., 2004; Yoon & Kim, 2004; Kwun & Kim, 2010; Noh et al., 2016). The PCR method, using several primers, was applied to the identification of three endemic Spanish barbel species: Barbus bocagei, B. graellsii and B. sclateri (Callejas & Ochando, 1998). Results indicated that Barbus bocagei and B. graellsii were more closely related to each other than they were to B. sclateri. In this study, PCR analysis has revealed a significant genetic distance between two population/species couples. PCR enabled to detect the existence of population/species discrimination and genetic variation in two geographic populations of tonguesole. The hierarchical polar dendrogram indicates two main cluster 1 (GUNSAN 01–GUNSAN 11) and cluster 2 (ATLANTIC 12–ATLANTIC 22) from two geographic tonguesole populations.
The shortest genetic distance displaying significant molecular difference was between individuals’ GUNSAN no. 02 and GUNSAN no. 01 (genetic distance=0.038). In the long run, individual no. 02 of the ATLANTIC tonguesole was most distantly related to GUNSAN no. 06 (genetic distance=0.958). These results demonstrate that the Gunsan tonguesole population is genetically different from the Atlantic tonguesole population. Phylogenetic relationships among 5 Haliotis species and one hybrid were conducted by calculation of the distance coefficient and construction of a phylogenetic tree based on PCR data (Kim et al., 2000). These branched off into two clusters: cluster I was formed by H. discus hannai, H. discus, H. gigantea, H. sieboldii, and the hybrid, which was subsequently re-divided into two sub-clusters. As mentioned above, the potential of PCR analysis to identify diagnostic markers for the identification of two tonguesole populations has been demonstrated. In teleost, a phylogenetic tree was constructed using UPGMA cluster analysis based on a total of 3,744 distinguishable fragments in gynogenetic clones from the silver crucian carp, Carassius auratus gibelio Block (Zhou et al., 2000). The inter-population similarity indices and genetic distance values indicated that the Jamuna-Padma population pair of the Indian major carp (Catla catla) was genetically closer than the Halda-Jamuna and the Halda-Padma population pairs, which agreed with the geographical distances between them (Islam et al., 2005). The two phylogenetic trees resulting from the neighbor-joining and parsimony analyses showed the same topology in distinguishing the Mullidae species (Mamuris et al., 1999). This confirms that the method is a suitable tool for DNA comparisons, both within and between individuals, species, and populations. As a rule, using various oligonucleotides primers, this PCR method has been applied to identify specific markers specific to species and geographical population, as well as genetic diversity/polymorphism in diverse species of organisms (McCormack et al., 2000; Yoon & Kim, 2003a; Kim et al., 2004; Kwun & Kim, 2010; Noh et al., 2016).