INTRODUCTION
At birth, mammalian ovary has fixed number of primordial follicles which are arrested at the first meiotic prophase stage. At puberty, several primordial follicles are selected to start folliculogenesis again. Granulosa cells (GCs) and CCs which are somatic cells that surround oocyte start to differentiate during folliculogenesis. These follicular cells play the important roles in providing nutritious factors needed for oocyte maturation. Oocytes also modulate follicular development by transporting many factors to these somatic cells. Therefore, proliferation of CCs and the interactions between oocyte and follicular cells are important to COC development and in determining oocyte quality.
Mammalian oocytes are arrested germinal vesicle (GV) stage so called prophase I and resume meiosis with disappearance of nuclear membrane, namely germinal vesicle breakdown (GVBD). Condensed homologous chromosomes are separated by MI spindles and asymmetric division occurs with involvement of many genes essential to support meiotic maturation (Mehlmann, 2005; Kim et al., 2012). After extrusion of polar body, oocytes are arrested again at meiotic metaphase II (MII) stage before fertilization occurs (Sagata, 1996), and the penetration of sperm lead oocytes to end up the meiotic division.
As a specific inhibitor of RNA polymerase II, α-amanitin is involved in the synthesis of mRNAs (Lindell et al., 1970; Sekeris & Schmid, 1972). It is a cyclic peptide that interrupts translocation of RNA polymerase II during transcription process. Many transcriptional assays have been performed by using α-amanitin experimentally. During in vitro maturation of oocytes, it affects bovine oocyte maturation and embryonic development (Kastrop et al., 1991; Memili & First, 1998). In addition, nuclear maturation was suppressed by α-amanitin in porcine COC (Meinecke & Meinecke-Tillmann, 1993).
Molecular mechanism of α-amanitin that affects in vitro maturation of COC is not well known. Therefore, the aims of the present study were 1) to investigate the rates of oocyte maturation and changes in COCs according to the treatment of α-amanitin during in vitro culture, and 2) to scrutinize the changes of specific gene expression in oocytes and CCs.
MATERIALS & METHODS
All ICR mice were obtained from Koatech (Pyeoungtack, Korea) were at the CHA Research Institute of CHA University to collect oocytes. All procedures described were reviewed and approved by the Institutional Animal Care and Use Committee of CHA University and were performed in accordance with the Guiding Principles for the Care and Use of Laboratory Animals.
To obtain oocytes, 3-week-old female ICR mice were injected with 5 IU PMSG and then sacrificed 46 h later. COCs were recovered from ovaries by puncturing the preovulatory follicles with 27-guage needles in M2 medium. M2 medium (Sigma, St. Louis, MO) containing 0.2 mM 3-isobutyl-1-methyl-xanthine (IBMX; Sigma) was used to inhibit GVBD during oocyte collection. CCs were removed from oocytes mechanically by repeated pipetting using a fine-bore glass pipette, collected and washed twice with 0.1% PVA/PBS.
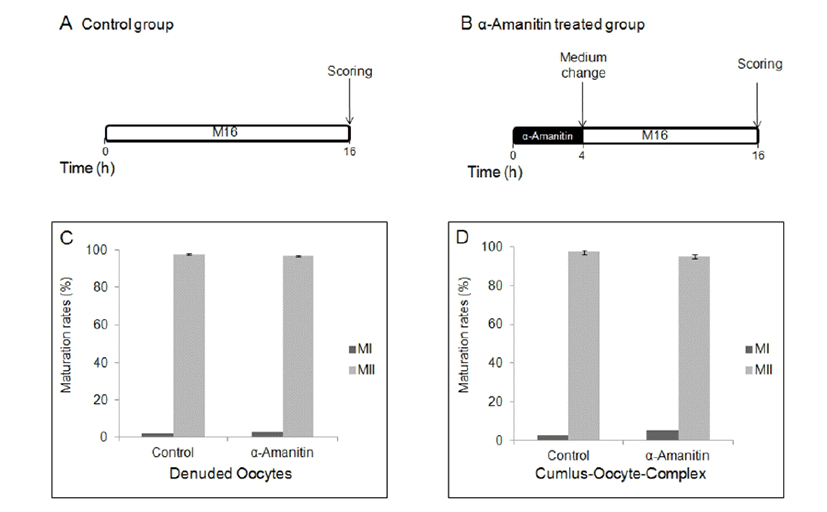
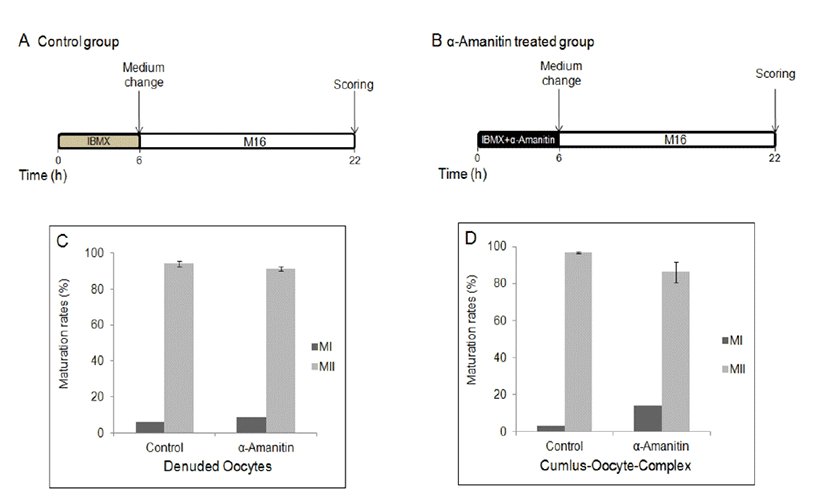
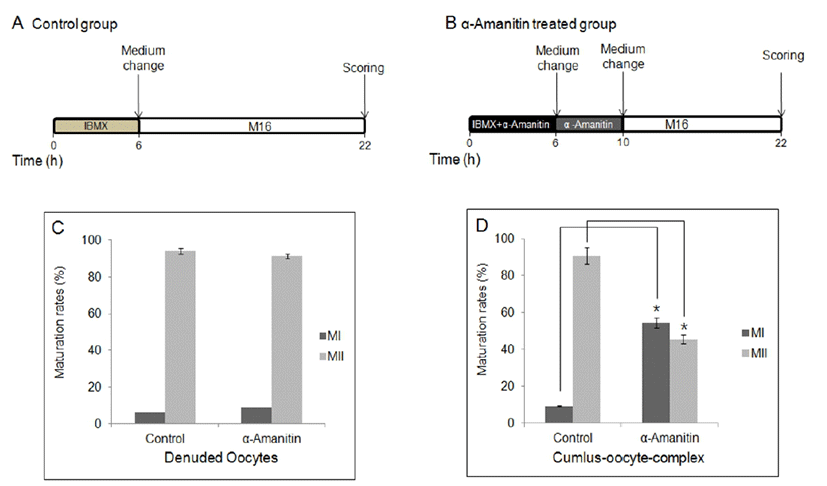
To determine the effects of α-amanitin to the oocyte maturation, α-amanitin was treated in culture medium at different period of time during in vitro culture. First, DOs and COCs were cultured in M16 medium supplemented with α-amanitin for 4 h followed by culture in the plain M16 medium for 12 h (Fig. 1A, B). Second, DOs and COCs were cultured in M16 medium supplemented with α-amanitin and 0.2 mM IBMX for 6 h followed by culture in the plain M16 medium for further 16 h (Fig. 2A, B). Finally, DOs and COCs were cultured in M16 medium supplemented with α-amanitin and 0.2 mM IBMX for 6 h followed by culture in M16 containing α-amanitin for 4 h and moved into the plain M16 medium for further 12 h (Fig. 3A, B). All cultures were accomplished in 5% CO2 at 37°C.
Messenger RNA was isolated from DOs and CCs using the Dynabeads mRNA DIRECT kit (Invitrogen Dynal, Oslo, Norway) according to the manufacturer’s instructions. Briefly, oocytes were resuspended in 300 μl lysis/binding buffer (100 mM Tris-HCl [pH 7.5], 500 mM LiCl, 10 mM EDTA, 1% LiDS, and 5 mM dithiothreitol [DTT]) for 5 min at room temperature. After vortexing, 20 μl prewashed Dynabeads oligo dT25 were mixed with cell lysates and annealed by rotating for 5 min at room temperature. The beads were separated with a Dynal MPC-S magnetic particle concentrator, and poly(A)+ RNAs were eluted by incubation in 14 μl Tris-HCl (10 mM Tris-HCl, pH 7.5) at 73°C for 2 min.
Complementary DNA was synthesized from mRNA or total RNA using 0.5 μg oligo dT primers according to the SuperScript Preamplification System protocol (Gibco-BRL). Polymerase chain reactions (20 μl) were carried out in 20 mM Tris-HCl (pH 8.4), 50 mM KCl, 1.5 mM MgCl2, 0.2 mM deoxynucleotide triphosphates (dNTPs), 25 pM of each primer, and 2.5 units Taq DNA polymerase (Promega, Madison, WI). Single oocyte-equivalent cDNAs were used as templates for PCR analysis. PCR products were separated by 1.5% agarose gel electrophoresis, followed by quantified using Image J software. The primer sequences and product sizes are summarized in Table 1. Relative gene expression levels were calculated after normalization to that of Gapdh. All experiments were repeated in triplicates.
| Gene | Accession no. | Primer sequence* | Annealing temperature (°C) | Product size (bp) |
|---|---|---|---|---|
| Ptx3 | NM_008987.3 | F : 5’-GCATCCTGTGAGACCAATGA-3’ R : 5’-GGGTTCCACTTTGTGCCATA-3’ |
60 | 111 |
| Has2 | NM_008216.3 | F : 5’-GCTGTACAATGCCATGTGGT-3’ R : 5’-GGATGACTGTGGCAATGAGA-3’ |
60 | 101 |
| Tnfaip6 | NM_009398.2 | F : 5’-TCCACGGCTTTGTAGGAAGA-3’ R : 5’-GACGGATGCATCACTCAGAA-3’ |
60 | 101 |
| Gja1 | NM_010288.3 | F : 5’- AAGAACACGGCAAGGTGAAG-3’ R : 5’- TACCACTGGATCAGCAGGAA-3’ |
60 | 106 |
| Gja3 | NM_001271623.1 NM_016975.3 |
F : 5’-TCATCTTCATGCTGGCTGTG-3’ R : 5’-GCATCTGGGTTGAAGTGGTT-3’ |
60 | 115 |
| Gja4 | NM_008120.3 | F : 5’-CAGCAACCTGAGCAAGAAGA-3’ R : 5’-CACAGCAGCCTTGAAGATGA-3’ |
60 | 73 |
| Gja10 | NM_010289.2 | F : 5’-TGCAAGAGAGAGTTGGGGTTT-3’ R : 5’-CACTATGCCGTTGTCCCTTT-3’ |
60 | 101 |
| Gjb1 | NM_008124.2 | F : 5’-AGAGACACAAGGTGCACATCTC-3’ R : 5’-CATGAAGACAGCCTCGAACA-3’ |
60 | 89 |
| Gjc1 | NM_001159382.1 NM_001159383.1 NM_008122.3 |
F : 5’-TGTGTGCAGCAGACTTCCTT-3’ R : 5’-CCCAAACCCTAAGTGAAGCA-3’ |
60 | 148 |
| FSHr | NM_013523.3 | F : 5’-TGTGGTCATCTGTGGTTGCT-3’ R : 5’-GCTTGGCAATCTTGGTGTCT-3’ |
60 | 92 |
| LHr | NM_013582.2 | F : 5’-AGTGGCCTTTGTCGTCATCT-3’ R : 5’-GCCATGCATGTGAAGTCTGT-3’ |
60 | 141 |
| Gdf9 | NM_008110.2 | F : 5’-TTGGCAGTCTCTTCAGTCCA-3’ R : 5’-GGGAGATCTTTCCACCTCAA-3’ |
60 | 106 |
| Prps1 | NM_021463.4 | F : 5’-TTGATATCCCGGTGGACAAT-3’ R : 5’-AGGGCCAGAAAAGATTCCAT-3’ |
60 | 356 |
| Rpe | NM_025683.2 | F : 5’-CTTGCCATCAAACCAGGAAC-3’ R : 5’-GCACTGCCAGACACAATCAT-3’ |
60 | 248 |
| Rpia | NM_009075.2 | F : 5’-TGCAGCGAATAGCTGAAAGA-3’ R : 5’-ACAGCCATTCGAAGTTCCAC-3’ |
60 | 394 |
| Taldo1 | NM_011528.4 | F : 5’-TGACGCTCATCTCTCCCTTT-3’ R : 5’-GCCAGCTTGCTGTTATCCTT-3’ |
60 | 365 |
| Tkt | NM_009388.5 | F : 5’-TCGGAGCTCTTCAAAAAGGA-3’ R : 5’-TCTCTGTTGCAACTCCATCG-3’ |
60 | 325 |
| H1foo | NM_008872 | F: 5’-GCGAAACCGAAAGAGGTCAGAA-3’ R: 5’-TGGAGGAGGTCTTGGGAAGTAA-3’ |
60 | 378 |
| Gapdh | BC092294 | F: 5’-ACCACAGTCCATGCCATCAC-3’ R: 5’-TCCACCACCCTGTTGCTGTA-3’ |
60 | 451 |
RESULTS
When DOs and COCs were cultured in M16 supplemented α-amanitin for 4 h followed by culture in plain medium for 12 h, oocytes in all groups developed to MII stage (Fig. 1B, C, D) with no difference in control oocytes matured in plain medium for 16 h (Fig. 1A). There was no difference between DOs and COCs (Fig. 1C, D). When DOs and COCs were cultured in IBMX-supplemented medium with or without α-amanitin for 6 h followed by culture in plain medium for 16 h (Fig. 2A, B), no change had occurred in any group of oocytes, denuded or cumulus-enclosed (Fig. 2C, D). However, meiotic maturation arrest has occurred when oocytes, especially in a form of COCs, were cultured in the medium with α-amanitin for 4 h during GVBD occurs after 6 h of pretreatment of α-amanitin in the IBMX-supplemented medium (Fig. 3B, D). Most oocytes from COCs were arrested at MI stage (54.52%). In contrast, maturation of DOs was not affected by any culture conditions at all (Fig. 1C, 2C, and 3C).
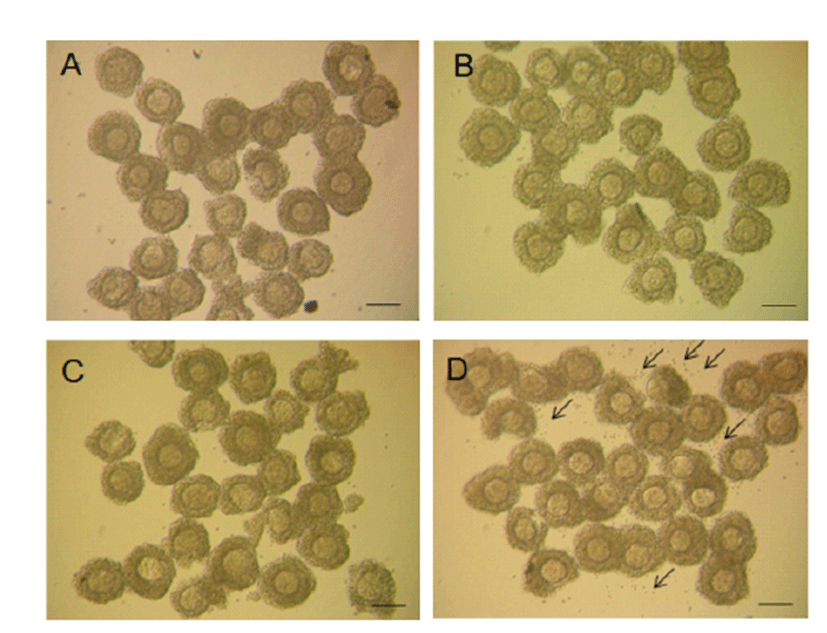
As depicted in Fig. 4, morphological changes, such as cumulus expansion derived by α-amanitin treatment was not vivid except the more scattered CCs found around the COCs in α-amanitin treated group (Fig. 4D). We focused on this feature as an abnormal maturation of COCs and tried to find causes of scattering of CCs in terms of cumulus expansion and intercellular junction formation. As cumulus expansion makes CCs be isolated from the mass, we investigated changes in gene expression of cumulus expansion markers (Has2, Ptx3 and Tnfaip6) using RT-PCR (Fig. 5). Interestingly, the expression of cumulus expansion markers decreased in α-amanitin treated CCs but not in control CCs.
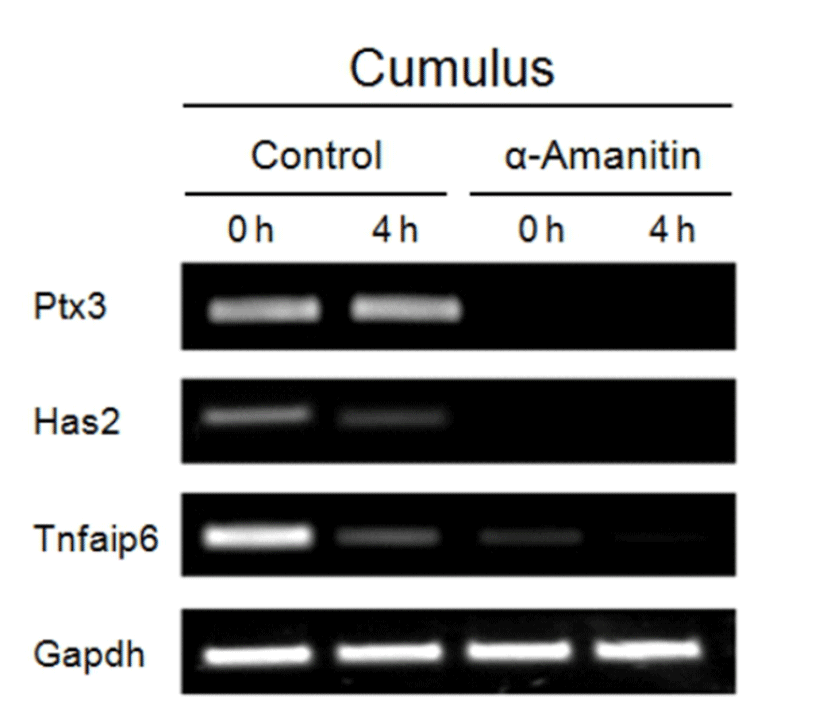
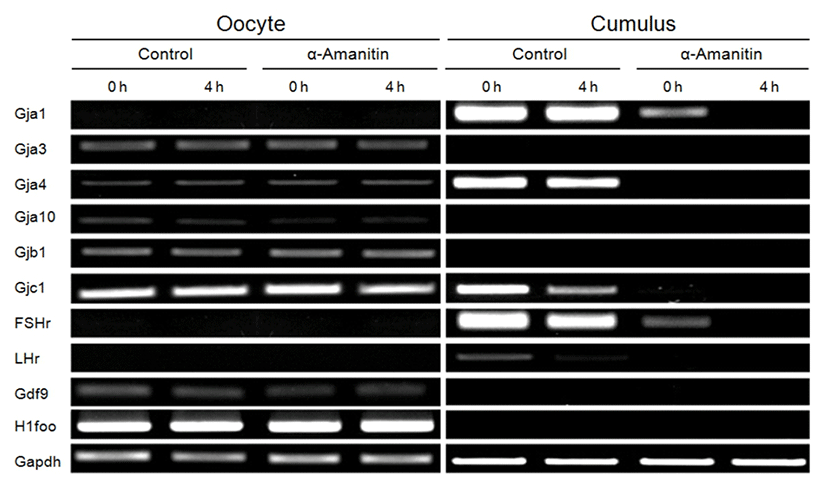
Intercellular connections among CCs’ networking are mediated by gap junctions. We measured expression of gap junctional genes (Gja1, Gja3, Gja4, Gja10, Gjb1, and Gjc1; Fig. 6). Although expression of gap junctional genes did not change in oocytes in both control and α-amanitin treated groups (Fig. 6, left panel), expression of gap junctional genes (Gja1, Gja4 and Gjc1) and gonadotropin receptor genes (FSHr and LHr) was dramatically decreased in α-amanitin treated CCs (Fig. 6, right panel). Expression of Gja3, Gjb1 and Gja10 was not observed in CCs both in control and α-amanitin group. Gdf9 and H1foo as well as gonadotropin receptors were used as an internal marker for cleanly and completely separated collection of oocytes and CCs, respectively.
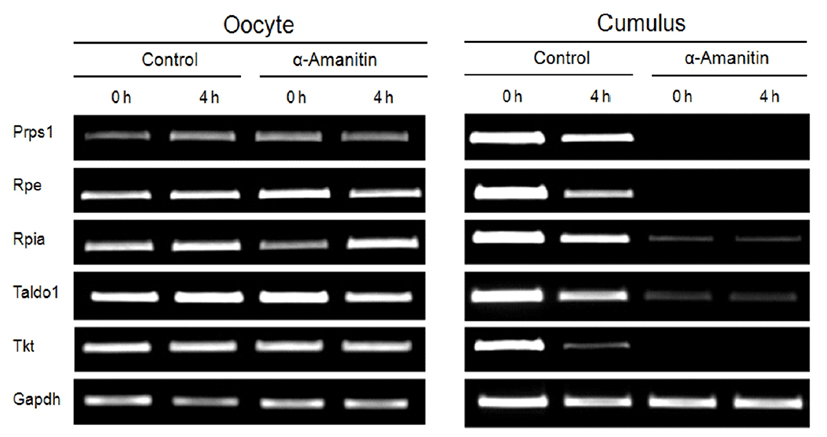
During in vitro maturation of COCs, normal metabolic capacities of CCs are essential to support normal meiotic maturation of the oocytes. Thus, we investigated changes in the transcription level of enzymes related to the cellular metabolism (Prps1, Rpe, Rpia, Taldo1, and Tkt) by α-amanitin treatment (Fig. 7). Oocytes cultured with or without α-amanitin showed no changes in the expression of metabolic genes (Fig. 7, left panel). However, their expressions in CCs disappeared dramatically in α-amanitin treatment group (Fig. 7, right panel) suggesting changes in the metabolism of the CCs, but not oocytes, has occurred by α-amanitin treatment.
DISCUSSION
In the present study, we demonstrated that the molecular mechanism of effects of α-amanitin on the in vitro maturation of COCs. Meiotic maturation of oocytes from the α-amanitin-treated COCs was arrested at MI stage when COCs were exposed to the α-amanitin during GVBD period (4 h) with pretreatment at GV stage for 6 h, while maturation of DOs was not affected. It was interesting to find out that the gene expression was not affected in oocytes itself, but in CCs. Therefore, we clearly showed that the transcription in CCs but not oocytes was affected by α-amanitin, and healthy and normal CCs itself and interactions between oocytes and CCs are extremely important to the development and maturation of the oocytes. We also demonstrated that α-amanitin affected the expression of genes for gap junctional proteins and metabolic enzymes, as well as for cumulus expansion. Decrease of these genes’ expression is related with the featured CCs spreading.
Gap junction is a intercellular connection between cells that directly connects cytoplasm of cells and it enables small materials such as ions, metabolites, and second messengers to move between cells (Bruzzone et al., 1996). It is formed by a combination of the complex of connexin (CX) proteins. Open gap junction consists of six connexins and each protein is four-pass transmembrane protein (Unger et al., 1999). There are many types of connexin protein and feature of gap junction is cell-specific and is related to what type of connexins combined to form gap junctions. In the mammalian ovary, expressions of Connexin 26 (CX26), Connexin 30.3 (CX30.3), Connexin 32 (CX32), Connexin 40 (CX40), Connexin 43 (CX43), and Connexin 45 (CX45), Connexin 5 (CX57) were identified (Kidder & Mhawi, 2002).
Especially, CX43 is involved in follicular development (Ackert et al., 2001; Wiesen & Midgley, 1993). Oocyte-specific depletion of CX43 resulted in infertility (Gershon et al., 2008). CX37 forms gap junctions between oocytes and GCs, and the folliculogenesis in the CX37 null mouse was arrested at the antral stage (Simon et al., 1997). Therefore, abnormal gap junctions lead to the abnormalities in cellular growth and development.
Since gap junctions are unstable proteins, enough mRNA levels should be maintained for proper regeneration (Laird, 1996). In this study, CCs’ transcriptional activities for gap junctional genes (Gja1, Gja3, Gja4, and Gjc1) were affected by α-amanitin treatment suggesting that there should be abnormalities in gap junctions and connections among the CCs as well as between CCs and oocytes. Therefore, these problems in cell-to-cell communications in COCs may lead unfavorable environment for oocytes development and maturation inside of the COCs. We also found decreased expression of cumulus expansion marker genes (Ptx3, Has2, and Tnfaip6) and speculated that it may be related with the modifications in gap junctions.
Cellular metabolism of CCs is important part of supporting in vitro maturation of COCs, and we found that the α-amanitin-treated CCs showed decreased expression of enzymes involved in the regulation of metabolism. Especially, these enzymes are important for generation of nucleic acid and energy sources. Prps1 is related to nucleotide synthesis (Kim et al., 2007). Rpe is a ribulose-5-phosphate-3-epimerase involved in pentose-phosphate pathway (Spencer & Hopkinson, 1980). Rpia is an important enzyme involved in the reversible conversion between ribose-5-phosphate and ribulose-5-phosphate in the pentose-phosphate pathway. Taldo1 is important in providing ribose-5-phosphate for nucleic acid and lipid synthesis. Its mutation causes transaldolase deficiency and early infantile liver failure (Balasubramaniam et al., 2011). Tkt is important gene essential for embryonic development, energy generation and nucleic acid synthesis. It regulates meiotic cell cycle through pentose phosphate pathway in oocytes (Kim et al., 2012). As α-amanitin blocked transcriptional activities of CCs, metabolic capacities were diminished. Therefore the reduced supply of nutrients and other small molecules vital for oocyte maturity was suggested and it may lead difficulties in developing oocyte normally to the MII stage in vitro.
Based on the results of the present study, we determined that the transcriptional activities of oocytes were not shown during in vitro maturation, and concluded that the transcriptional activities in CCs during in vitro maturation of COCs are essential for maintaining their gap junctions among cells and their metabolic capacities for successful maturation of oocytes.