
INTRODUCTION
Human mesenchymal stem cells (MSCs) can be isolated from a variety of tissues, such as bone marrow, adipose tissue or other tissues. They are promising sources for cell based therapy because of the potential to differentiate into various cell lineages including adipocytes, osteoblasts and chondrocytes (Galderisi & Giordano, 2014). It has been expected that MSCs will replace damaged tissue and/or induce endogenous repair after homing to the tissue of injury following transplantation in vivo. Compared to other tissues, human adipose tissue is an abundant and easily accessible for the isolation of stem cells. Hence adipose-derived stem cells (ADSCs) have been recognized as a suitable cell source for tissue engineering and regenerative medicine applications (Chen et al., 2013). ADSCs have been examined for their clinical potentials to treat disease such as graft versus host disease (Le Blanc et al., 2008), cartilage damage (Bhumiratana et al., 2014), myocardial infarction (Jiang et al., 2014), liver malfunction (Li et al., 2013) and osteogenesis imperfect (Pereira et al., 2014).
Previously, we observed that human eyelid adiposederived stem cells could form aggregation during highdensity culture in the presence of human serum (HS) in vitro (Song et al., 2012). Since it is well known that FBS alone does not induce an aggregation of plastic-adherent MSCs in vitro, it was questioned whether the aggregation phenomenon might be due to the presence of cell-aggregating factor(s) in HS or due to the absence of the factor in FBS. The present study aimed to give answers to these questions. Furthermore, to unveil the nature of the aggregation phenomenon, it needs to examine whether the aggregation could form only by HS or by other animal sera or serum components. For the purposes, we examined if human follicular fluid, cord blood serum, or animal serum and components could induce the aggregation of MSCs.
Both differentiation of adult stem cells and their maintenance by self-renewal are regulated by intrinsic and extrinsic signals. In recent years, many of the signaling pathways and transcriptional mechanisms that underlie stem cell fate decisions and the establishment of differrentiated traits have been elucidated. Extrinsic cues, such as cell-cell and cell-matrix adhesion, as well as a wide range of growth factors and small molecules, have been shown to affect stem cell behavior (Brizzi et al., 2012). It has been indicated that adherent junctions and gap junctions are essential for the promotion of MSC stemness and proliferation (Wong et al., 2008). For example, MSCs are known to form functional gap junctions with adult cardiac myocytes both in vitro and in vivo (Valiunas et al., 2004; Hahn et al., 2008). Tight junctions (TJs) are specialized plasma membrane microdomains which are highly enriched in cholesterol that encircle cells at the apical end of the lateral membrane are aligned at the apical side of adherent junctions (Yamada et al., 2013). Claudin-6, one of essential component of TJs, has been shown as a cell surface marker of hESCs and hiPSCs due by their specific expression (Ben-David et al., 2013). Present researches focus on the possible involvement and elucidation of the roles of TJs in the aggregation phenomenon of the human adult stem cells during high-density culture.
MATERIALS AND METHODS
HS (Millipore, Temecula, CA) was heat-activated at 56°C for 30 min and kept frozen at –20°C until use. Cord blood serum (CBS) and human follicular fluid (HFF) were kindly donated from CHA Gangnam Medical Center in Seoul. Porcine serum (PS, Sigma), sheep serum (SS, Millipore), bovine serum (BS, Gibco, Grand Island, NY) were used after treatment like HS. Unless indicated elsewhere, all reagents were purchased from Sigma (Seoul, Korea).
Human abdominal adipose tissues were obtained from the patients undergoing liposuction with informed consent. All experiments were approved by Institutional Review Board of Seoul Women’s University. After removal of red blood cells by several washing with Dulbecco’s phosphatebuffer saline (DPBS, Gibco), adipose tissue was mixed with 1x volume of 0.15% type I collagenase (Gibco) and incubated at 37°C for 1 h with gentle shaking. Then 1x volume of culture medium consisting of DMEM-low glucose type (DMEM-LG; Gibco), 10% FBS (Gibco), 100 U/mL penicillin, 0.1 mg/mL streptomycin, and 3.7 mg/mL sodium bicarbonate was added and centrifuged at 3,000 rpm for 10 min. Cell pellets were washed twice, and then plated in 25-cm2 culture flask (Nunc, Roskilde, Denmark) containing 5 mL of the culture medium. Cells were cultivated at 37°C with 5% CO2. When adherent cells were 70~80% confluent, they were detached using Hank’s balanced salt solution containing 0.125% trypsin (Gibco) and 1 mM EDTA (trypsin/EDTA) at 37°C for 2 min and re-suspended in the culture medium at a density of 4,000 cells/cm2. The culture medium was changed twice a week. ADSCs between passage (p) 2 and p6 were used throughout this study.
ADSCs were seeded in 6-well or 12-well plate (Nunc, Jiangsu, China) at a density of 2.8×104 cells/cm2. Cells were cultivated with DMEM-LG containing 5% or 10% of FBS, HS, CBS, HFF, PS, SS or 1% of bovine serum albumin (BSA). Sometimes cells were seeded in 24-well dish (Nunc) at same density. After culture in HS, aggregates of ADSCs (HS-Agg) were isolated using a pipette tip. HStreated but non-aggregated cells (HS-Ex) were harvested after detachment using trypsin/EDTA solution. Collected cells were kept at –20°C before use.
Total RNA was isolated using Tri-reagent (Ambion, Carlsbad, CA) according to the manufacturer’s instructions. RT-PCR was carried out using a Technic PCR system TC- 100 (Bibby Scientific, Staffordshire, UK). Total 7.5 μg (1 μg/μL) of RNA was reverse-transcribed using the following RT mixture: 20 mM MgCl2 (Bio Basic, Ontario, Canada), 10 × PCR buffer (Bio Basic), 10 mM dNTPs mixture (Bio Basic), 0.5 mg/mL oligo (d)T15 (Bionics, Korea) for RTPCR or 0.5 mg/mL random hexamer (Roche, Branchburg, NJ) for qRT-PCR , 40 U/μL RNase inhibitor (Bio Basic) and 200 U/μL AMV-RT (Invitrogen, Carlsbad, CA). RT reaction was allowed to occur for 60 min at 37°C. PCR was performed using the following mixture: 25 mM MgCl2, 10 × PCR buffer, 5 U/μL Taq polymerase, 2.5 mM dNTP, 10 μM sense and antisense primer. Amplification was performed for 35 cycles at a denaturing temperature of 94°C for 30 sec and an extension temperature of 72°C for 30 sec. Annealing temperature was set depending on the species of primer. The PCR products electrophoretically separated on agarose gels were analyzed using an image analyzer (ULTima, Hoefer, Leicestershire, UK). qRT-PCR was performed using a buffer containing 1 μL of template cDNA, 10 μL SYBR Green I Master (Roche, Mannheim, Germany), and 10 pM of each primer using Light Cycler 480 Real-Time System (Roche). Relative expression levels of cDNA were normalized to the expression of 18s rRNA using the comparative CT (2-ΔΔCT) method. Primers used in this study are shown in Table 1. PCR mixture components were purchased from Takara (Shiga, Japan).
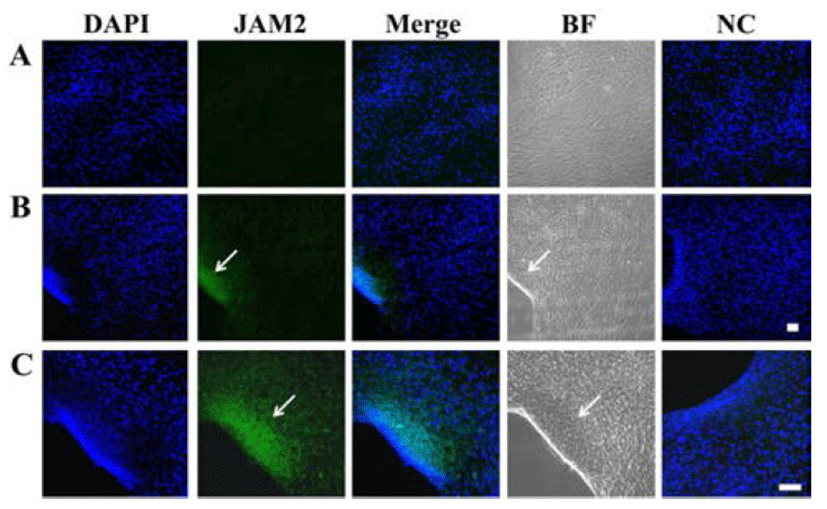
Cell cultures on 8-well slides were fixed in 4% paraformaldehyde for 1 h at 4°C. They were washed with washing buffer (0.1% BSA in DPBS) and incubated in a blocking solution consisting of 2% BSA in PBS for 1 h at room temperature with gentle shaking. Then cells were incubated with rabbit polyclonal anti-JAM2 antibody (1:400) (Abcam, Cambridge, UK) for overnight at 4°C with gentle shaking. They were then incubated with the goat antirabbit IgG-FITC (1:100) (Santa Cruz, CA) for 1 h. After washing, cell nuclei were stained with 1 μg/mL DAPI in 0.1 M PBS for 2 min. Fluorescent images were visualized by confocal microscopy (C1+, Nikon, Melville, NY) or fluorescence microscope (Axioskop2+, Carl Zeiss, Thornwood, NY).
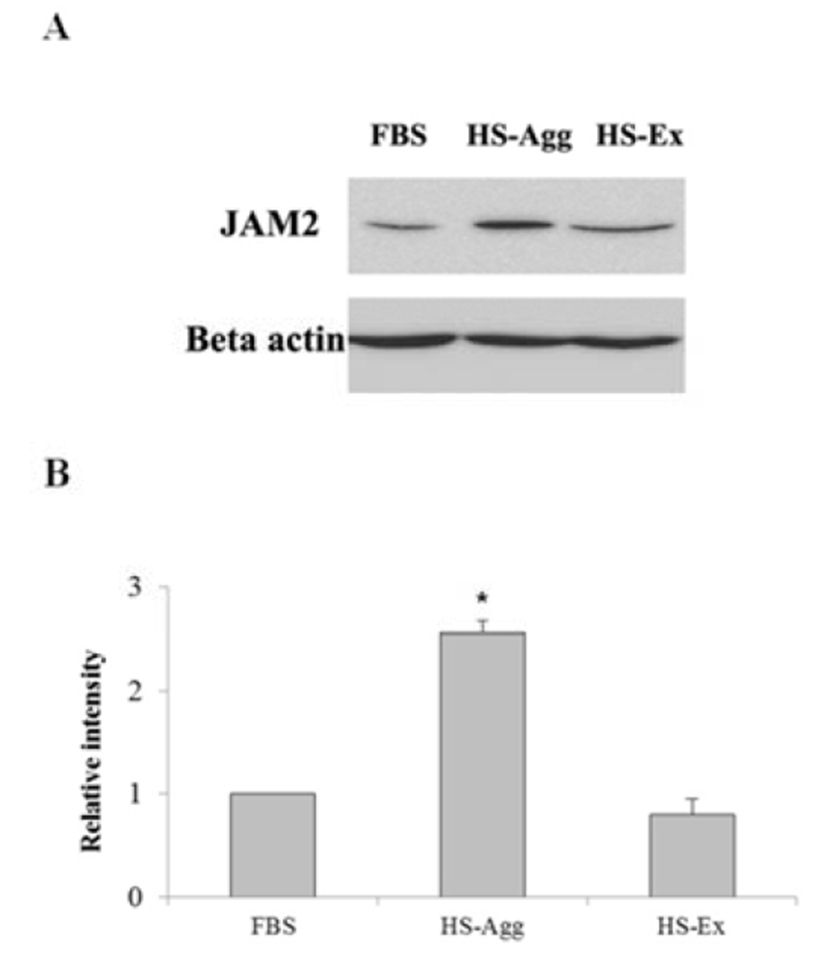
Cells were homogenized in lysis buffer consisting of 50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 1% sodium deoxycholate, 1% Triton X-100, 1% sodium dodecyl sulfate (SDS), and 25x protease inhibitor (Roche, Mannheim, Germany) for 30 min on ice. Protein sample was mixed with 2× sample buffer (12.5 mM Tris-HCl (pH 6.8), 4% SDS, 10% 2-mercaptoethanol, 20% glycerol, 0.004% bromophenol blue) and heated at 90°C for 5 min. Following separation by 10% SDS polyacrylamide gel electrophoresis (SDS-PAGE), proteins on the gels were electrically transferred to nitrocellulose membranes (Millipore, Bedford, MA). Then membranes were incubated with the rabbit polyclonal anti-JAM2 antibody or mouse monoclonal antibody against β-actin (1:800, Santa Cruz) in blocking buffer (4.9% BSA in washing buffer) for overnight at 4°C. Then the membrane was incubated in HRP-conjugated goat anti-mouse IgG for 1 h at 37°C. Membranes were the visualized by ECL (BioFX, Aden Prairie, MN). Data were analyzed using ImageJ program (NIH, Bethesda, MD).
RESULTS
During culture of ADSCs in the presence of 10% FBS, none of the cells from three different lines showed any aggregation or morphological change in every experiment. However, same cells but cultivated in HS exhibited aggregation, some of which were located in the center of wells whereas others were formed in the vicinity of the wells (Fig. 1A). Cell aggregation was observed within 5 days from the beginning of culture (Fig. 1B). However, when cells were pre-treated with FBS for 1 day before replacing with HS-medium or BSA-medium, the aggregation formation was delayed to 8-10 days. To examine whether FBS could inhibit the HS-induced aggregation, cells were cultivated in the mixture of FBS and HS. In the presence of 10% mixture, most wells exhibited aggregation-positive within 12 days. In the presence of 5% mixture, only 60% of wells showed aggregation-positive after 15 days without further aggregation (Fig. 1C). In contrast, all wells containing cells with either 5% or 10% HS exhibited aggregationpositive within 8 days. These results demonstrate that FBS could not inhibit the aggregation of ADSCs induced by HS, implying that cell aggregation might be induced by certain component(s) present in the HS.
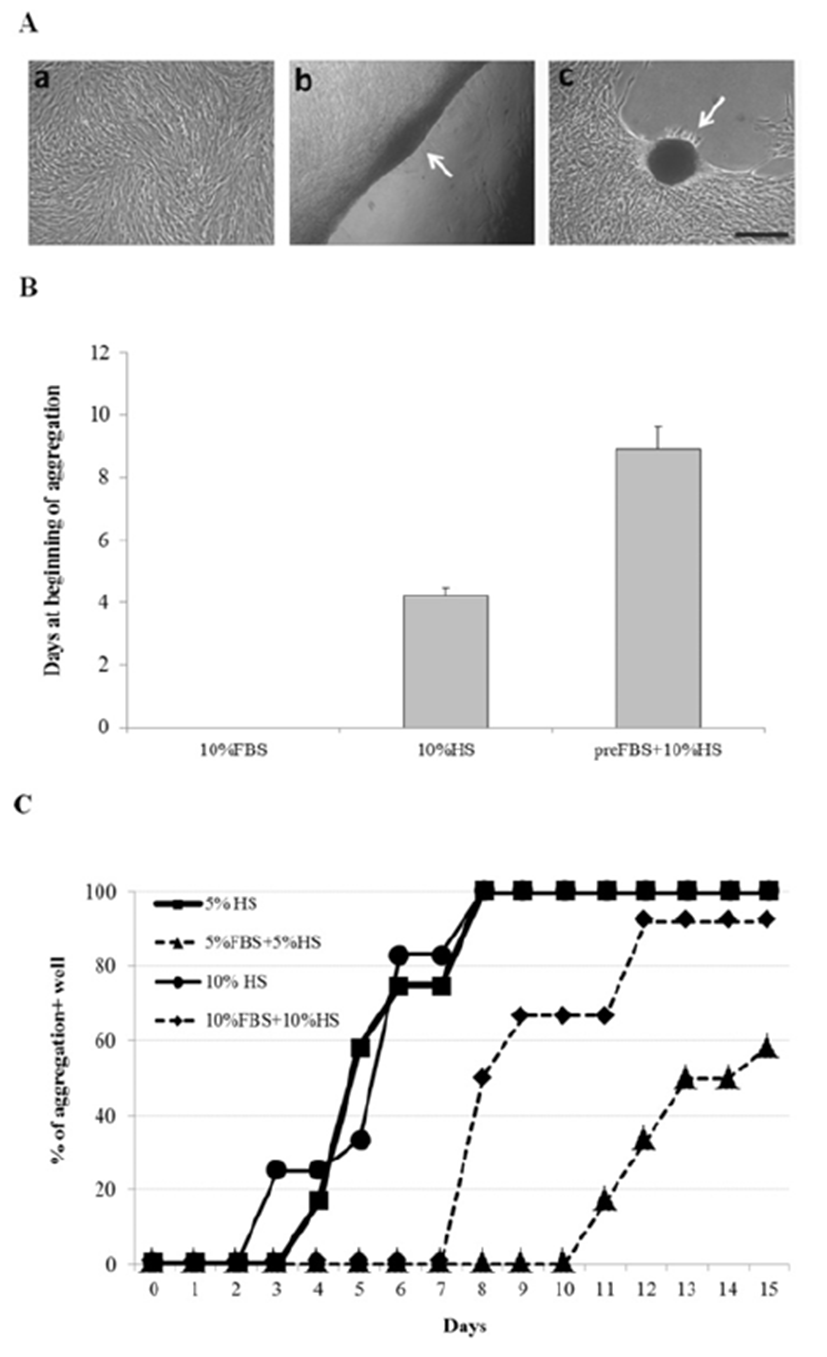
It was further examined whether other human body fluids could induce similar aggregation of ADSCs. All culture wells containing either CBS or HFF showed the appearance of cell aggregation (Fig. 2A). However, while HS showed 100% of aggregation-positive wells within 4 days of culture, CBS or HFF induced 100% or 80% aggregation 6 or 8 days after culture, respectively. Pre-treatment of ADSCs with FBS delayed the aggregation by 6 days during the subsequent culture in HFF whereas pre-treatment with FBS resulted in little difference between HS-culture and CBS-culture (Fig. 2B). These results demonstrate that human body fluids such as CBS and HFF might contain same or similar cell-aggregating factors as HS, while the aggregating activity of HFF seemed weaker than other two.
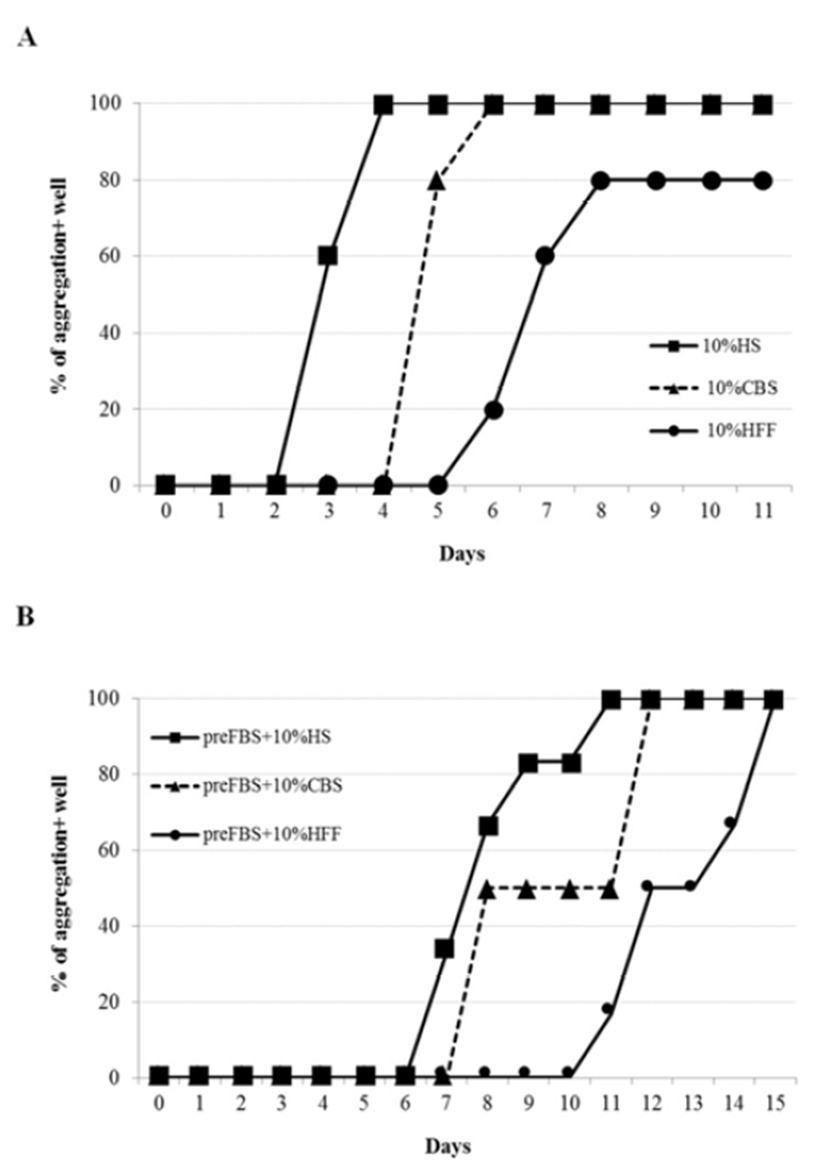
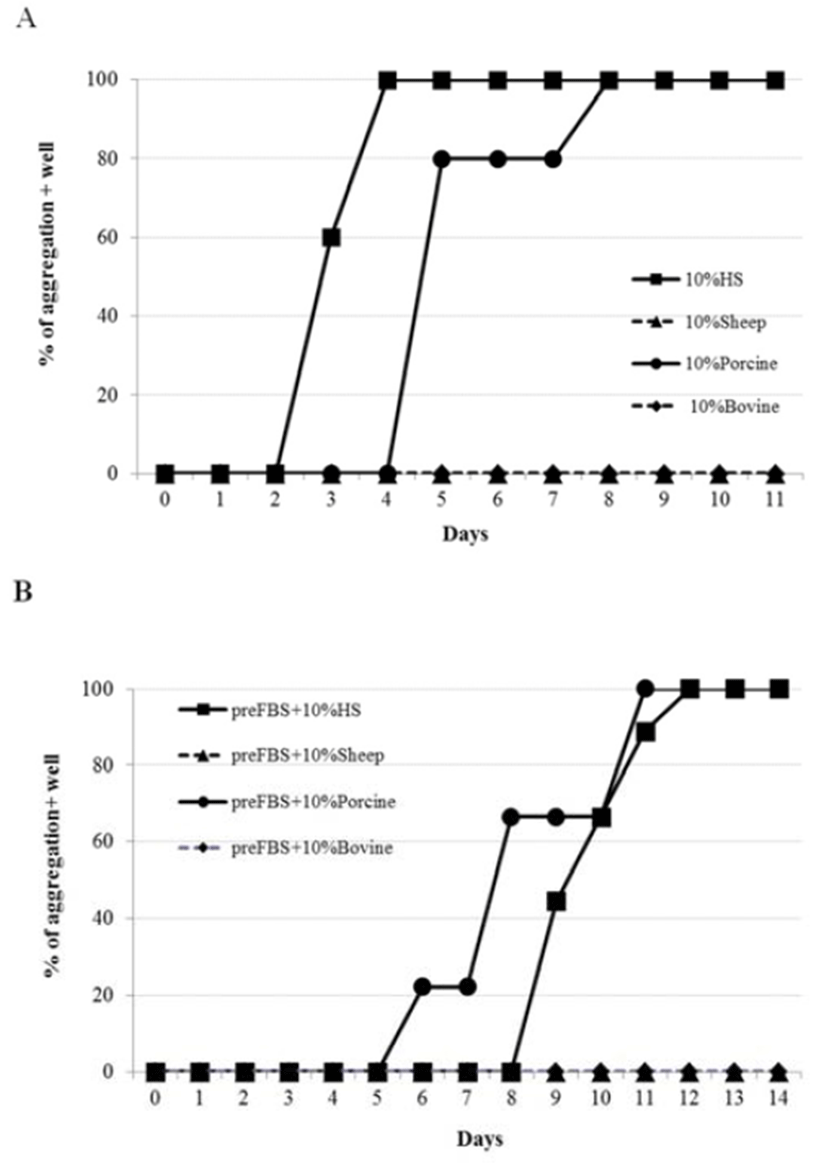
To see if animal serum other than HS could induce similar aggregation of ADSCs, effects of sheep serum (SS), porcine serum (PS) and bovine serum (BS) were examined. Similar to HS, PS-medium induced distinctive aggregation (Fig. 3A). However, the aggregation seemed to take place 2 days later compared to the HS-medium. In contrast, neither SS nor BS showed any appearance of aggregation at all under the same culture condition. When cells were pre-treated with FBS, again it delayed the aggregation. However, there was little difference in the culture periods required for the 100% aggregation-positive wells between HS- and PS-treated groups (Fig. 3B). These results showed that cell aggregating activity by body fluids is not humanspecific, rather is depending on the animal species.
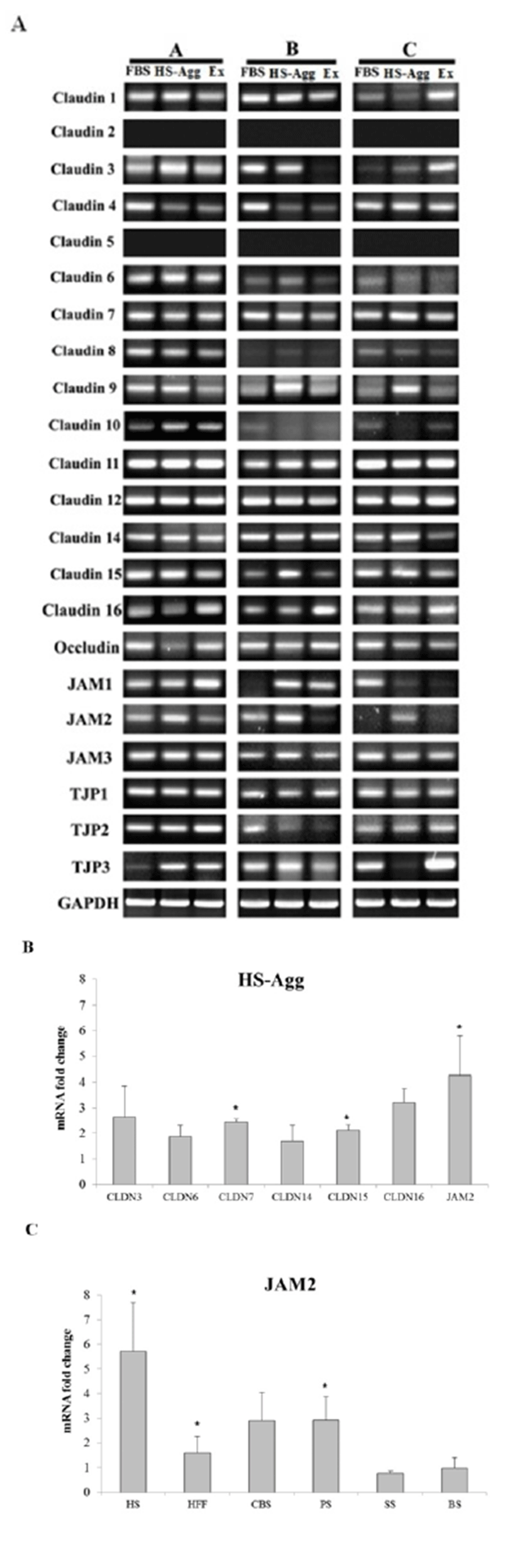
aggregated cells maintained tightly adhering structures during culture for many days, it was questioned what kinds of cell adhesion molecules might be involved in the rigid aggregation. As shown in Fig. 4A, conven-tional RT-PCR analyses showed that ADSCs consistently expressed mRNAs of CLDN7, CLDN11, CLDN12, CLDN14, TJP1, and JAM3 genes. In contrast, cells inconsistently expressed genes of CLDN3, CLDN4, CLDN6, CLDN8, CLDN9, CLDN10, CLDN16, OCLN, JAM1, TJP2, and TJP3, depending on cell lines. They never expressed CLDN2 and CLDN5 genes. Quantitative RT-PCR analyses using three different cell lines showed greater expression of CLDN3, -6, -7, -15, -16 genes and JAM2 gene in HS-Agg than cells cultivated with FBS or HS-Ex (Fig. 4B). Of these mRNAs, mRNA of JAM2 showed the greatest expression. Compared to the JAM2 mRNA expression in cells cultivated with FBS, more than five times greater expression was observed in HS-Agg. Cells cultivated with HFF, CBS or PS also showed greater expression than cells with FBS. In contrast, cells with SS-culture or BS-culture showed similar expression level of JAM2 mRNA as cells cultivated with FBS (Fig. 4C).
Immunocytochemical staining showed that human JAM2 protein was distinctly expressed in ADSCs that has begun the aggregation during HS-culture, whereas none of the cells cultivated in FBS exhibited staining (Fig. 5). Western blot analysis demonstrated that JAM2 protein was expressed in cells cultivated with FBS or HS (Fig. 6A). However, the staining intensity of the protein was most prominent in HS-Agg compared to FBS group or HS-Ex group (Fig. 6B). Taken together, these results suggest that HS treatment could induce an aggregation in ADSCs, and only cells undergoing aggregation appear to enhance the JAM2 protein expression unlike remaining cells which do not undergo aggregation during the HS treatment.
DISCUSSION
The present study demonstrates that the aggregation property of ADSCs during high-density culture could be dependent on the property of serum components replacing FBS, and that JAM2 molecule, one of TJs family, might play a role in the aggregation.
Most cell culture technique utilizes FBS as a source of nutritional and growth factors, which supports prerequisite condition for most types of cell growth. However, various human or animal body fluids have also been shown adequate or beneficial to human cell culture. For example, HS and CBS have been shown to support proliferation and differentiation of MSCs which are intended for cell-based therapy (Aldahmash et al., 2011; Wang et al., 2012). Previously we have observed that human eyelid adiposederived stem cells could aggregate during high-density culture in the presence of HS (Song et al., 2012). In the present study using ADSCs, we observed that, in addition to HS, CBS and HFF could also induce the aggregation. These results lead to suggest that cell aggregation might be a generalized phenomenon which can take place in other adult stem cells. We also found that the aggregation is not unique to HS treatment, rather is general as seen in cells cultivated with CBS or HFF. Moreover, porcine serum produced morphologically similar aggregation. Considering that bovine and sheep sera, unlike porcine serum, failed to induce any aggregation, our data suggest that the aggregation phenomenon might be dependent on specific components of serum or body fluid. Regarding to the presence of the aggregation-inducing factor(s), we observed that HS could induce the aggregation even in the presence of FBS which does not induce any cell aggregation. These findings indicate that cell aggregation could be induced by the presence of inducible factors in human body fluids or porcine serum. Absence of aggregation in cells cultivated with FBS, bovine serum or sheep serum is due by the absence of aggregating factors, and not by the presence of inhibitor of aggregating factors. Yet there is no known candidate molecule responsible for the aggregation.
Cell-cell communication is necessary for the interaction between cells and is essential for cells to make an adjustment of their physiological event. Recently, studies have demonstrated that integrin-extracellular matrix (ECM) interaction induces and/or influence hMSC differentiation (Frith et al., 2012). The formation of cell-cell contact is carried out by junctional complex. One of the junctional complex consists of TJs family members, one group of which, known as JAMs have been shown to play a major role in supporting tight cell-cell adhesion (Garrido-Urbani et al., 2014). It regulates paracellular permeability and associates with the actin cytoskeleton, formation and maturation of cell-cell contact. In the present study, qRT-PCR analyses demonstrated that, compared to ADSCs cultivated with FBS, cells cultivated with HS exhibited higher level of mRNA expression of CLDN3, -6, -7, -15, and -16 genes among the TJs members. Among these, particularly, JAM2 mRNA showed the highest increase. Regarding that this increased expression of JAM2 mRNA in HS-Agg might somehow relate with aggregation, we have examined the expression level in cells which aggregated or not under various condition. Interestingly, when cells were cultivated in aggregation conditions such as in the presence of HFF, CBS or PS, and then examined, cells of all conditions exhibited greater expression than cells cultivated FBS. In contrast, when cells were cultivated in non-aggregation conditions such as in the presence of sheep serum or bovine serum, and then examined, none of them showed difference from FBS-cultured cells. Furthermore, JAM2 protein expression were markedly increased in HS-Agg compared to those cells cultivated with FBS and cells of HS-Ex. These results suggest that JAM2 molecule might play an important role in the aggregation of ADSCs which is induced by specific serum or body fluids. In support of this hypothesis, results of immunocytochemical studies indicate the localization of JAM2 protein was restricted to the cells undergoing aggregation, and no or little protein was seen in cells with FBS-culture or cell of HS-Ex. Moreover, western blot results also demonstrate that JAM2 protein expression was most prominent in HS-Agg than cells with FBS-culture or HS-Ex.
TJs family include transmembrane proteins such as claudin, occludin, and junctional adhesion molecules (JAM), the latter which are associated with claudin through their cytoplasmic protein, ZO-1 (Bazzoni et al., 2003). JAM family is composed of JAM1, JAM2, JAM3, and JAM4/ JAML, with each have different functions. Of these, JAM2 plays a role in many events such as interaction between embryo and uterine tissue at the time of implantation (Su et al., 2012), melanoma cell metastasis (Ghislin et al., 2011) and the blood-testis barrier between Sertoli cells (Wang Y et al., 2009). These reports indicate a role of JAM2 in cellcell adhesion events. In addition, it plays a crucial role in the adjustment of transendothelial migration and mediated rolling of T lymphocytes dependent on integrin alpha4 and beta1 (Ludwig et al., 2009). Previously, GDF-5 has been shown to enhance the chondrogenic differentiation of human MSCs in vitro as revealed by increased expression of glycosaminoglycan and type II collagen (Zhang et al., 2011). Later study demonstrates that the enhancing effect of GDF-5 might be mediated by the expression of connexin43, which appear to play an important role in in vitro chondrogenesis (Sun et al., 2012). Taken together, it is suggested that tight junction molecules such as JAM and connexin43 plays a role in cell migration as well as cellcell interaction. Present study showed that JAM2 molecule is involved in the aggregation of adult stem cells induced by specific serum or body fluid. Since ADSCs must migrate directing to an area where they aggregate, it is possible that JAM2 molecule in the present study might play dual roles, both migration and adhesion of ADSCs. One important thing is that while our study shows ADSCs responded to HS, CBS, HFF or PS resulting in the localized cell aggregation, only a small portion of cells exhibited dynamic response. Most cells remain unchanged and keep their position and shape. These observations imply that only a portion of cells is capable of responding to the signal. Further studies are needed to characterize these specific cells.
In summary, present study demonstrates that aggregation property of ADSCs during high-density culture might be dependent on yet unidentified components which are present in specific serum or body fluid, and that JAM2 molecule might play an important role in these specific components-induced aggregation of human adult stem cells in vitro.