
INTRODUCTION
Most fishes have a distinct spawning season and their annual spawning cycle is influenced by various environmental factors, especially photoperiod and water temperature (Biswas et al., 2004; Suquet et al., 2005). These environmental factors affect the Brain-Pituitary-Gonad (BPG) axis, and induce gonadal development and spawning through regulation by neuropeptide and sexual hormone in reproductive endocrine system (Shimizu, 2003; Howell et al., 2003; Levy et al., 2011; Shao et al., 2013). There are many studies of the influences of reproductive activity in fish, particularly the effects of photoperiod and water temperature on gonadal development.
Damselfish, Chromis notata is widely distributed in tropical and temperate waters and one of the important fishery species in Korea. The spawning of damselfish C. notata inhabiting Jeju Island, Korea occurred from June to August (Lee & Lee, 1987). In the temperate fish, photoperiod is the main proximal factor in the timing of gonadal maturation. Photoperiod alternations are used to stimulate or delay gonadal development (Falcon et al, 2003; Sarkar & Upadhyay, 2011). However, it is unclear whether damselfish utilize these environmental factors for initiation and termination of their reproductive activity. Therefore, the present study investigated the effects of photoperiod on gonadal activity of the damselfish in Jeju Island, South Korea. In addition, RT-PCR assays mRNA expression of GtH subunits (FSHβ, LHβ) in the pituitary gland under the artificial condition where photoperiod is controlled.
Materials and method
Sexually mature male and female damselfish Chromis notata (standard length 7.0 to 9.0 cm, body weight 12.0 to 30.0 g) were collected in coastal waters of the Bomok, Seogwipo, Jeju, South Korea. Adult animals were kept at the Marine Science Institute, Jeju National University, Jeju, South Korea, in 1.5 ton aquaria with an open seawater circulation system. The experimental fish were divided into three groups; 14L:10D, 12L: 12D and 10L:14D. They were reared for approximately 60 days from June to August 2011. Ten to fifteen fish from each group were randomly selected to compare gonadal development and GtHs expression under different conditions. The experimental fish were anesthetized with 2-phenoxyethanol (Sigma-Aldrich, St. Louis, Mo), then hypothalamus, pituitary and ovary were extracted. The extracted ovary of each specimen were weighted for calculation of the gonadosomatic index (GSI=gonad weight/body weight×100). For the histological analysis, ovaries samples were fixed in Bouin’s solution, embedded in paraffin, sectioned 5 μm thickness and stained with haematoxylineosin. The hypothalamus and pituitary sample were kept at – 80℃ until total RNA extraction.
The total amount of RNA from the hypothalamus and pituitary samples was extracted using RNAiso Reagent (TaKaRa Bio, Kyoto, Japan). The total RNA was treated with RQ RNase-free DNase (Promega, Madison, WI, USA) to prevent genomic DNA contamination. The hypothalamus and purity of the total extracted RNA from the pituitary was examined with the ratio of 1.7 and 2.1 for A260/A280. From 1 µg of the extracted total RNA, the reverse transcription reaction was performed to synthesize cDNA using the Transcriptor First strand cDNA Synthesis kit (Roche Diagnostics, Mannheim, Germany).
Primers for the experiments were produced by FSHβ and LHβ of damselfish C. notata that are listed on NCBI. A qPCR was conducted with 20 ng of cDNA using SYBR green premix PCR kit (Takara) in CFX96TM Real-time System (Bio-Rad, Hercules, CA, USA). PCR was performed at 95℃ after the initial denaturation. Afterwards, the PCR reaction was performed by 40 cycle of denaturation for 45 s at 94℃, annealing for 45 s at 58℃, and extension for 1 min at 72℃. Expression of the FSHβ and LHβ genes in each sample was normalized to the amount of the internal control β-actin gene.
Results and Discussion
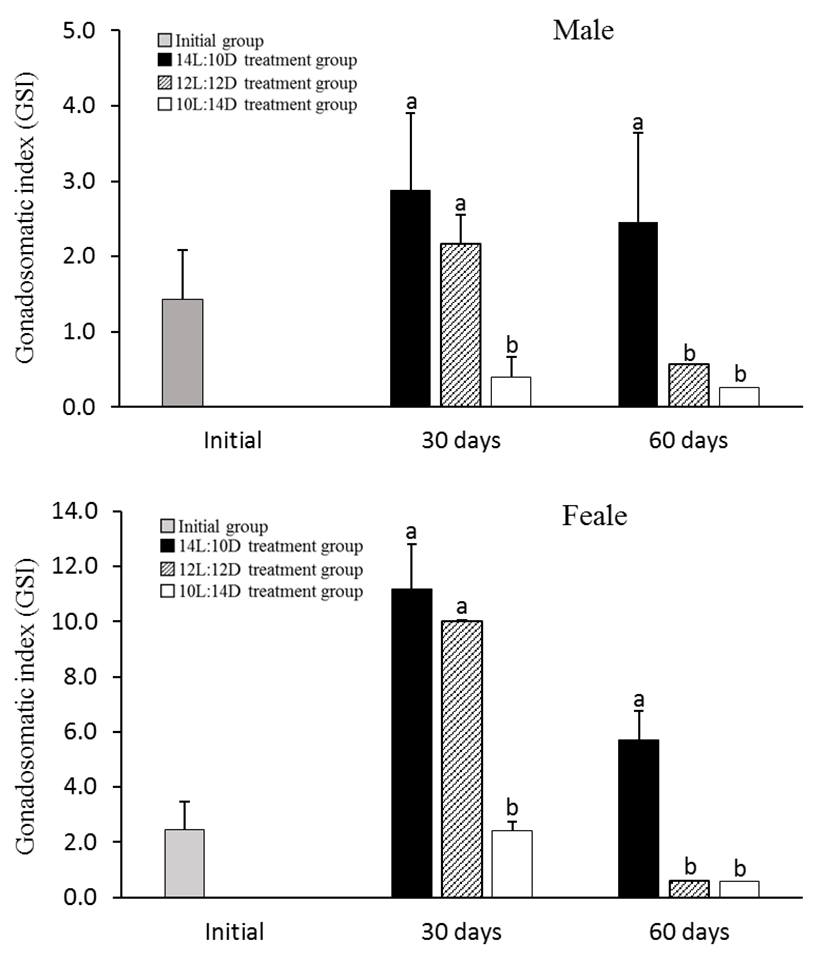
In 30th June 2011, the GSI of male and female in initial group was 1.43 ± 0.65 and 2.44 ± 1.05 respectively. However, in 30th July 2011, GSI of 10L:14D treatment group showed drastically decreased at 0.39 ± 0.29 of male and 2.40 ± 2.25 of female, showing a significant statistical difference (P<0.05). And in 30th August 2011, GSI of 12L:10D treatment group also showed drastically decreased at 0.56 ± 0.38 of male and 0.60 ± 0.05 of female but GSI of 14L:10D treatment group showed high levels at 2.46 ± 1.19 of male and 5.72 ± 1.02 of female (Fig. 1).
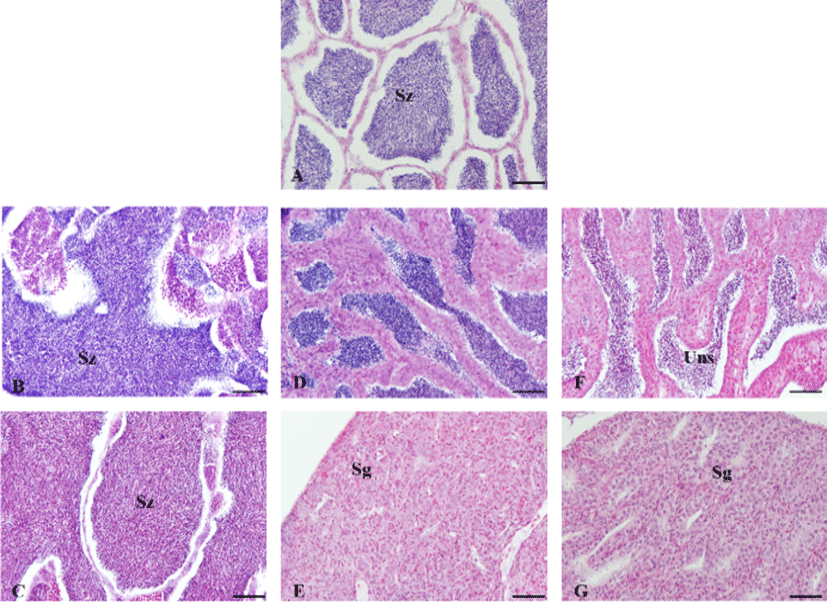
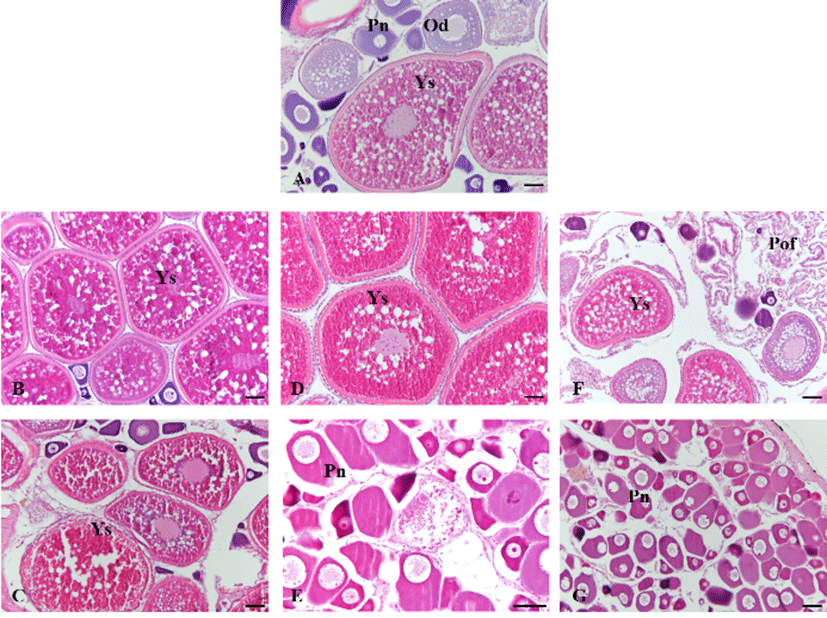
In 30th June 2011, the testis of initial group was mature phase, spermatozoa were filled in testicular lobules. After 30 days, the testis of 10L:14D treatment group was spent phase and observed undischarged sperm in the center of the testicular lobules but the other treatment group was mature phase. After 60 days, the testis of 10L:14D and 12L: 12D treatment groups were resting phase with spermatogonia but the testis of 14L:10D treatment group was still ripe phase (Fig. 2). In 30th June 2011, the ovary of initial group was mature phase with vitellogenic oocytes of 400 to 500 μm in diameter. After 30 days, the ovary of 10L: 14D treatment group was spawning phase and observed postovulatory follicle but the other treatment group was mature phase. After 60 days, the ovary of 10L:14D and 12L: 12D treatment groups showed early growing phase and contained mainly perinucleolus stage oocytes but the ovary of 14L:10D treatment group was still ripe phase, contained mature oocytes (Fig. 3).
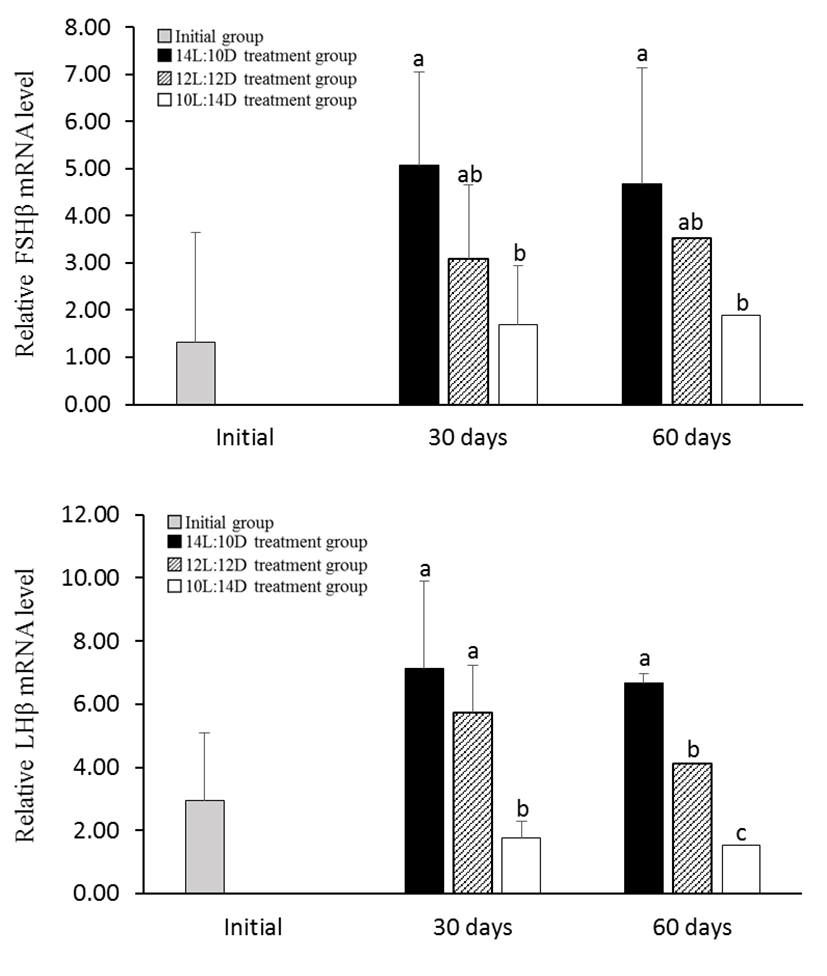
In the 10L:14D treatment group of male, the expression levels of FSHβ and LHβ mRNA was significantly decreased at July and August 2011 than the other treatment group. However 14L:10D treatment group showed high levels throughout all of the experiments (Fig. 4). Female GtH showed similar to male pattern. The expression levels of FSHβ and LHβ mRNA in the 10L:14D treatment group was significantly decreased at July and August 2011 (Fig. 5).
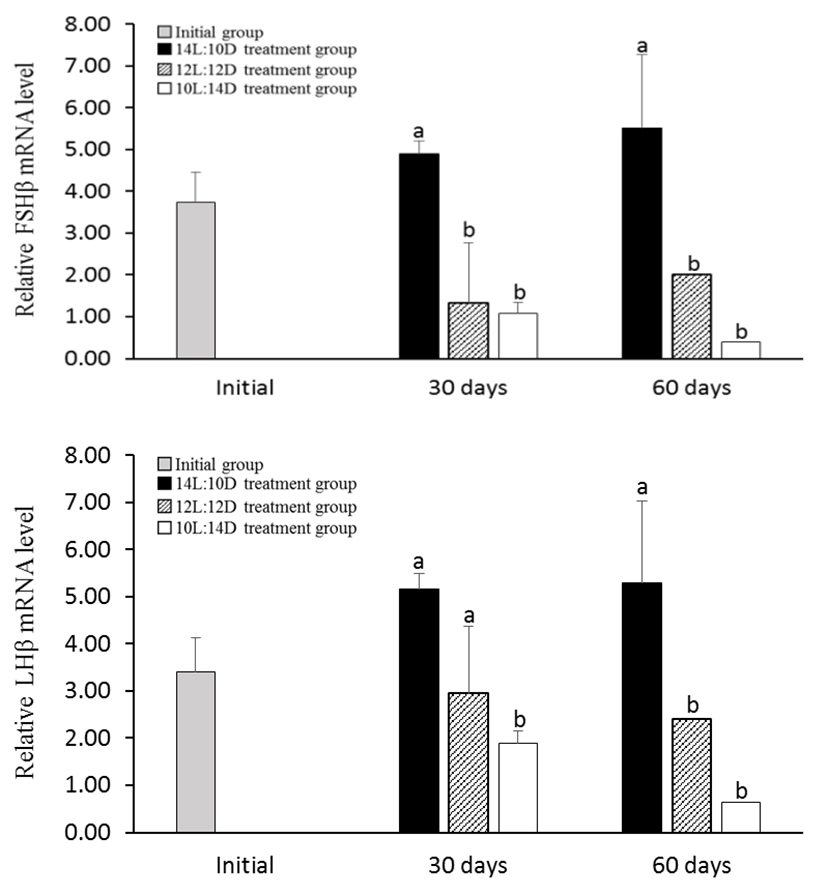
Spawning cycle and spawning timing of fish is determined by environmental conditions (photoperiod, water temperature and nutrient state etc.). Previous studies have shown effects of the environment, especially photoperiod and temperature, on sexual maturation and spawning, sexual maturation is regulated through the brain-pituitary-gonad (BPG) axis (Huber & Bengtson, 1999; García-López et al., 2006; Migaud et al., 2010). In the Nile tilapia, Oreochromis niloticus L., gonadal development was suppressed by treatment of constant and long photoperiod (Rad et al., 2006). In the sea bass Dicentrarchus labras, photoperiodic manipulation delayed maturation for three months with respect to controls (Zanuy et al., 1986). The Arctic charr, Salvelinus alpinus under decreasing photoperiod (16L:8D reducing to 8L:16D) delaying the reproductive cycle (reduction of up to 66% on sexual maturation of males and 32% females) (Duston et al., 2003). The Atlantic cod (Gadus morhua) under the 24L:0D photoperiod significantly decreased expression of FSHβ and LHβ during the sex maturation period (de Almeida et al., 2011). Amano et al. (1995), testing the two different conditions of short photoperiod (8L:16D) and long photoperiod (16L:8D), a shot photoperiod treatment induced sex maturation but long photoperiod treatment delayed sex maturation. Olive flounder, Paralichthys olivaceus, a marine species that spawns in the spring, also showed low GSI and delayed sexual maturation when treated under long photoperiod of 15L:9D from September to March, until the spawning season. Also, FSHβ and LHβ mRNA expression in pituitary decreased more in long photoperiod treatment group than in natural photoperiod (Kim et al., 2013). In this study, shot photoperiod treatment from July (spawning period; mature stage) was decreased gonadal development and reduced FSHβ and LHβ mRNA expression in pituitary of damselfish but long photoperiod treatment was maintained gonadal development and keeped FSHβ and LHβ mRNA expression in pituitary. These results show how the photoperiod affects the reproduction of several fish species, interfering with the control of gonadal maturation and pre-paration for reproduction. Photoperiod is considered to be the most effective environmental factor in controlling the reproductive cycle of damselfish.