
INTRODUCTION
Spontaneous abortion, also known as miscarriage is the natural death of an embryo or fetus before 20 weeks of pregnancy without surgical procedure for a human (Sierra & Stephenson, 2006). Several mechanisms have previously been described for pathogenesis of spontaneous abortion, including chromosomal anomalies, hormonal problems, uterine abnormalities, infections, and autoimmune disorders (Quenby et al., 2002). The mechanisms by which the fetus is protected from the maternal immune system during pregnancy are not fully understood. Tolerance of the fetus by the maternal immune system is thought to depend on the interactions of many cytokines produced by the uterus and transferred from maternal serum to the site of implantation.
In humans, the endometrium develops into decidua to be receptive the embryos during the implantation window under the influence of gestational hormones and cytokines produced by maternal tissues. Communication between trophoblastic cells in the embryos and decidual cells in the uterus is mediated by cytokines and cell surface receptors (Saini et al., 2011). Many cytokines are produced by trophoblastic and immune cells present within the decidua which include mainly T lymphocytes, macrophages and natural killer cells (van Mourik et al., 2009). Cytokines are commonly classified in one or the other category: interleukin-1 (IL-1), tumor necrosis factor (TNF), gamma-interferon (IFN-gamma), IL-12, IL-18 and granulocyte-macrophage colony stimulating factor (G-MCF) are well characterized as pro-inflammatory cytokines whereas IL4, IL-10, IL-13, IFN-α and transforming growth factor-β are recognized as anti-inflammatory cytokines (Corripio-Miyar et al., 2007; Hanada & Yoshimura, 2002). The net effect of an inflammatory response is determined by the balance between pro-inflammatory and anti-inflammatory cytokines. Pro-inflammatory cytokines are produced predominantly by activated macrophages and dendritic cells in the implantation sites and are involved in the up-regulation of inflammatory reactions. A high level of the pro-inflammatory T helper (Th)-1 and cytokines (IL-6, IL-8, and TNFα) characterizes early implantation (Mor & Koga, 2008; Yoshinaga, 2008). In addition, an array of cytokines in maternal serum which are transferred to the fetus may play an important role in implantation of embryo and maintenance of pregnancy.
Nesfatin-1 protein is originally known as a regulator of appetite and energy metabolism, which is expressed in hypothalamus (Oh-I et al., 2006; Shimizu et al., 2009; Stengel et al., 2009a,b; Stengel et al., 2010; Yosten et al., 2012). Recently, nesfatin-1/NUCB2 was expressed not only in the hypothalamus, but also in the peripheral organ such as digestive organs (Stengel et al., 2009a,b; Goebel et al., 2009; Xia et al., 2012), adipose tissues (Ramanjaneya et al., 2010), cardiac organ (Mimee et al., 2012), and reproductive organs (García-Galiano et al., 2010; García-Galiano et al., 2012; Gonzalez et al., 2012; Kim et al., 2011). Recently, it has been reported that nesfatin-1 expression levels are decreased significantly during pregnancy progresses in rat, suggesting that nesfatin-1 may play an important role during pregnancy and fetal development (Garces et al., 2014). We also previously demonstrated the expression of nesfatin-1/NUCB2 mRNA and protein in implantation sites during pregnancy (Chung et al., 2015).
However, it is not yet clear whether nesfatin-1/NUCB2 expressed in implantation site is associated with cytokines in maternal serum. Therefore, the purpose of this study was to examine the cytokine levels in maternal serum during spontaneous abortion using the CBA/j × DBA/2 mouse model and the relationship between nesfatin-1/NUCB2 expression in implantation site and maternal serum cytokine levels.
MATERIALS AND METHODS
CBA⁄j female mice, BALB⁄c male mice, and DBA⁄2 male mice were obtained from Koatech (Korea) 7 to 8 weeks of age and the mice were kept in cages. All cages were under controlled illumination (12:12 h light/dark cycle, lights on/off: 6 h/18 h) and temperature (22±2℃). Animals were fed a standard rodent diet and tap water ad libitum. When CBA⁄j female mice had reached 10 weeks of age, they were mated with DBA⁄2 or BALB/c males. CBA/j × DBA/2 mouse combination shows pregnancy loss as a model of spontaneous abortion, whereas CBA/j × BALB/c combination shows normal pregnancy. The morning sighting of a vaginal plug was considered day 0.5 of pregnancy. Pregnant female mice were sacrificed on day 14.5 of pregnancy under CO2 anesthesia. Blood samples were collected from each pregnant and non-pregnant mouse as a control by cardiac puncture. Uterine tissues of the implantation sites were quickly removed, and then stored at –70℃ for extracting total RNA preparation and protein extraction. Animal care and experimental procedures were approved by the Institutional Animal care and the use committee at the Seoul Women’s University in accordance with guidelines established by the Korea Food and Drug Administration.
Serum was obtained by centrifugation of blood samples at 6,000 rpm for 20 min. Mouse Cytokine Array Panels were used according the manufacturer’s instructions (ARY006, R&D Systems, Minneapolis, MN). Briefly, each sample was diluted and mixed with a cocktail of biotinylated detection antibodies and then incubated with Mouse Cytokine Array membrane. Therefore, any cytokine/ detection antibody complex being present had been bound by its cognate immobilized capture antibody on the membrane. After washing, Streptavidin-Horseradish Peroxidase and chemiluminescent detection reagents were added sequentially. Immunoreactivity was then visualized using enhanced chemiluminescence reagent. Densitometric analysis was then performed. The intensity was measured using image J software 1.45 (NIH, Bethesda, MD, USA). The following proteins were assessed: CXCL13, C5a, G-CSF, GM-CSF, CCL1, CCL11, sICAM-1, IFN-γ, IL-1α, IL-1β, IL-1ra, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-10, IL-13, IL-12p70, IL-16, IL-17, IL-23, IL-27, IP-10, CXCL11, KC, M-CSF, CCL2, CCL12, CXCL9, CCL3, CCL4, CXCL2, CCL5, CXCL12, CCL17, TIMP-1, TNF-α and TREM-1.
Uterine tissues of the implantation sites were homogenized with RNA isoplus (TaKaRa Bio, Shiga, Japan). After chloroform extraction and isopropyl alcohol precipitation, RNA was dissolved in RNase-free DEPC (TaKaRa Bio, Shiga, Japan) solution. The RNA concentrations were measured with the Nano-drop (Thermo Fisher Scientific Inc., Waltham, MA). First strand cDNA synthesis was performed using the extracted RNA and oligo dT, followed by double-strand synthesis in RT buffer (Invitrogen, Carlsbad, CA) with dNTP (BIO BASIC Inc., Ontario, Canada) and RTase (Invitrogen, Carlsbad, CA). qRT-PCR was performed in buffer solution containing template cDNA, SYBR Green (Roche, Manheim, Germany), and each primer. Primer pairs were as follows: NUCB2 forward 5-AAAACCTTGGCCTGTCTGAA-3; reverse 5-CATCGATAGGAACAGCTTCCA-3 and GAPDH forward 5-TTGATGGCAACAATCTCCAC-3; reverse 5-CGTCCCGTGACAAAA-TGGT-3 (BIONICS, Korea). The optimum temperature cycling protocol was determined to be 95℃ for 10 s, 60℃ for 10 s and 72℃ for 10 s using the Light Cycler 480 Real-time PCR System (Roche, Manheim, Germany).
Uterine tissues of the implantation sites were quickly removed and extracted the protein with EDTA homogenization buffer, the samples were SDS-PAGE and transferred to the PVDF membrane. The membrane was treated in a blocking solution and incubated with rabbit anti-rat nesfatin-1 antibody (H-003-22, Phoenix Pharmaceuticals) / anti-mouse β-actin antibody (sc-47778, Santa Cruz Biotechnology) followed by incubation with donkey anti-rabbit IgG-HRP (sc-2313, Santa Cruz Biotechnology) / donkey anti-mouse IgG-HRP (sc-2096, Santa Cruz Biotechnology), respectively. By Using the ECL Plus Western Blotting Detection Reagents (Amersham; GE Healthcare), the membrane was detected to investigate the expression level of nesfatin-1 protein.
RESULTS
Implantation sites were observed and counted in normal pregnant and abortion model mice. Total number of implantation sites was significantly increased in normal pregnant (10.15±0.58) compared to abortion model mice (7.1±0.60). Interestingly, the number of implantation sites was different between the left and right uterine horn in abortion model mice, but no difference between the left and right uterine horns of normal pregnant mice (Table 1).
| Normal pregnant (n=20) | Abortion model (n=20) | |
|---|---|---|
| Total | 10.15±0.58 | 7.1±0.60 |
| Left uterine | 4.8±0.28 | 4.7±0.27 |
| Right uterine | 5.35±0.30 | 2.4±0.33 |
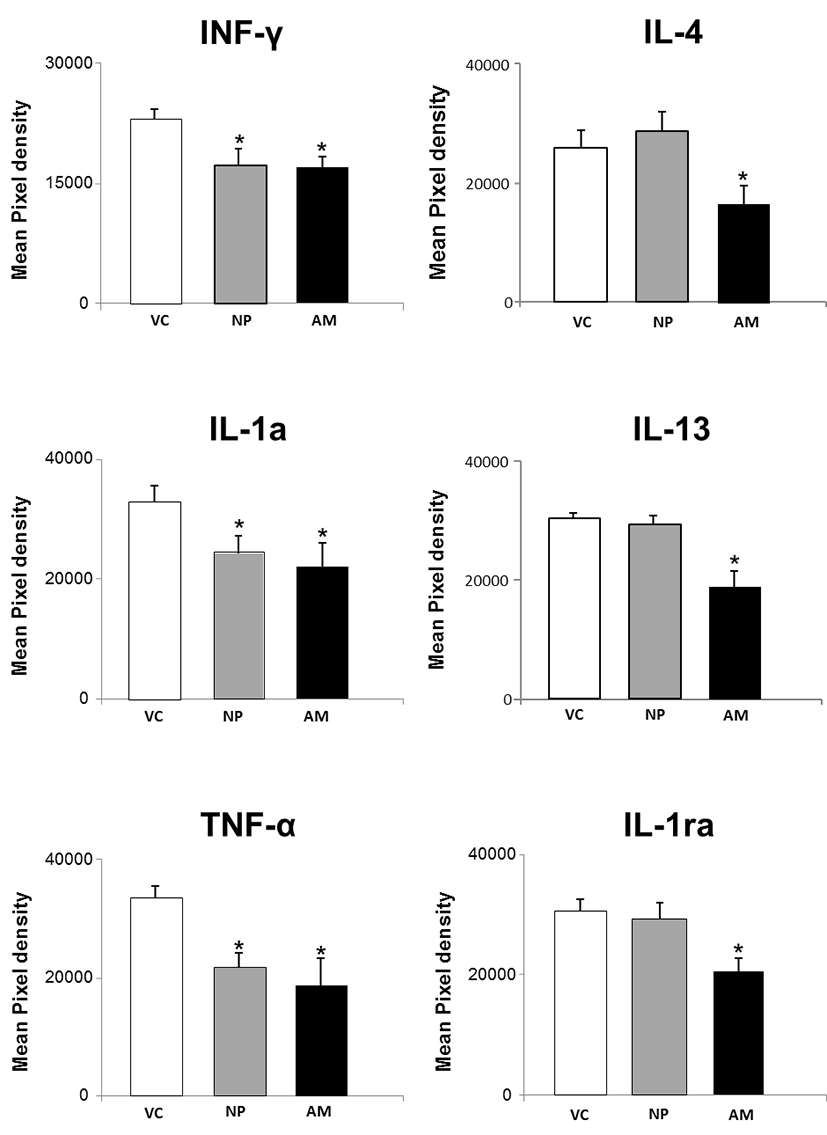
Serum cytokine levels of pregnant and non-pregnant mice were analyzed using cytokine array kit. The data are expressed as relative levels of selected cytokines based on the positive controls (Fig. 1). The expression of pro-inflammatory cytokines, including IFN-γ, IL-1α and TNF-α was significantly decreased in both normal pregnant and abortion model mice compared to non-pregnant controls. However, their levels were not different between normal pregnant and abortion model mice. On the other hand, the expression of anti-inflammatory cytokines, including IL-4, IL-13 and IL-1ra was not decreased in normal pregnant mice compared to non-pregnant controls. However, their levels were significantly decreased in abortion model mice compared to normal pregnant (Fig. 2).
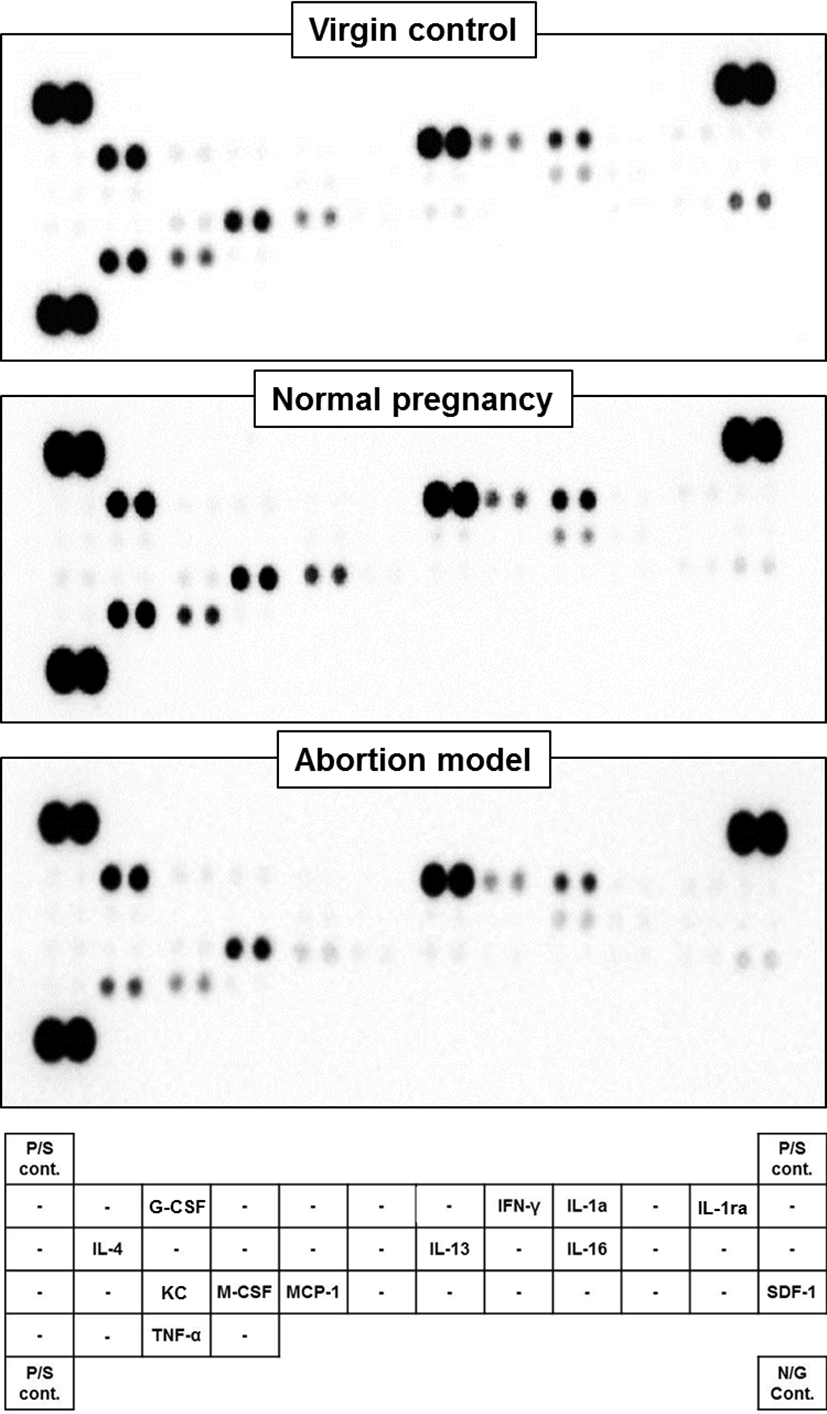
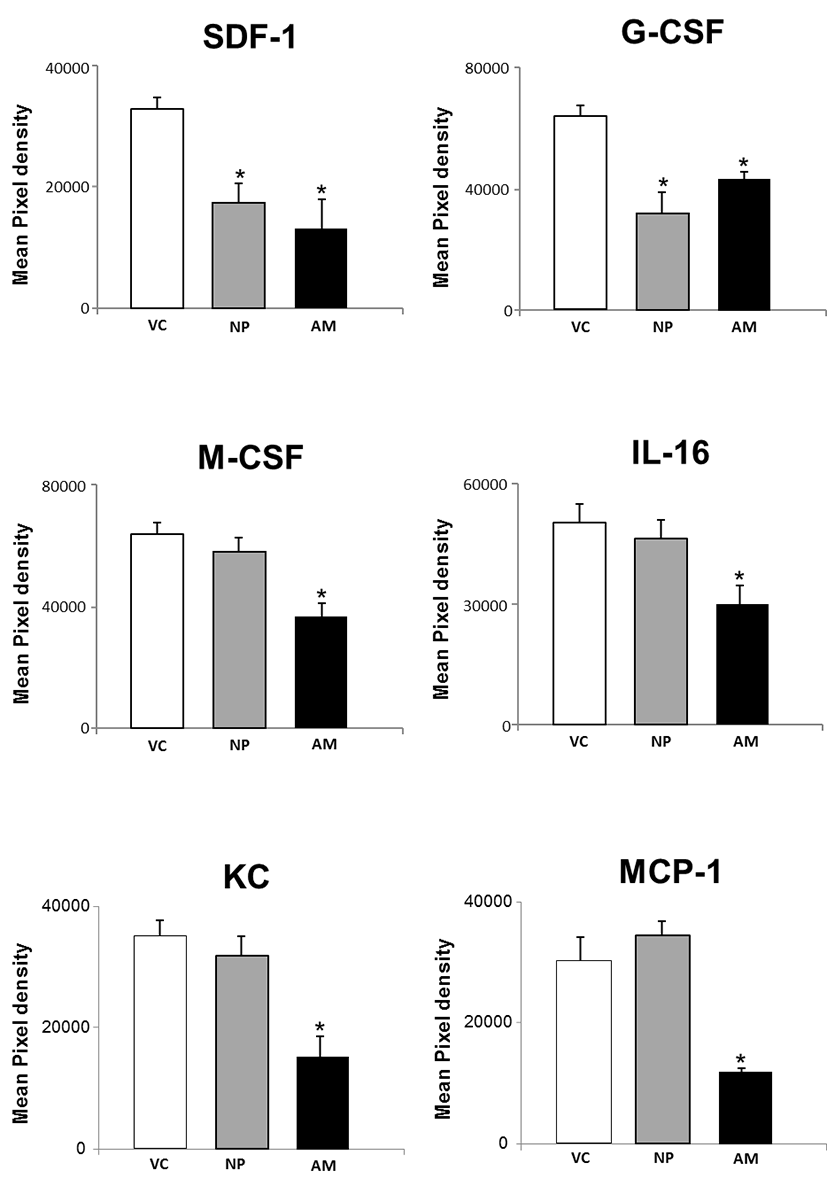
Next, we analyzed chemokines in the serum collected from normal pregnant and abortion model mice, and non-pregnant mice as a control. The expression of SDF-1 and G-CSF was significantly decreased in both normal pregnant and abortion model mice compared to non-pregnant controls, but showing no difference between normal pregnant and abortion model mice. On the other hand, the expression of M-CSF, IL-16, KC and MCP-1 was not decreased in normal pregnant mice compared to non-pregnant controls. However, their levels were significantly decreased in abortion model mice compared to normal pregnant (Fig. 3).
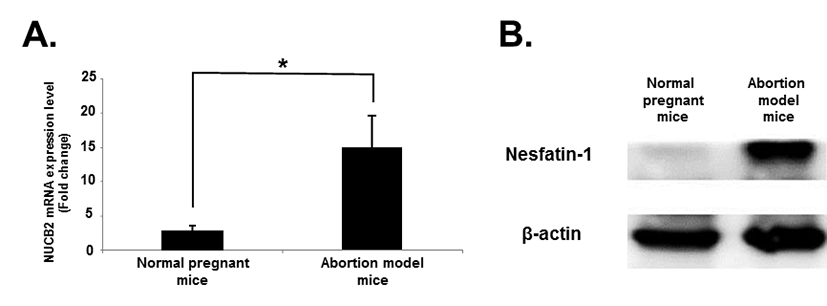
The expression levels of nesfatin-1/NUCB2 mRNA in the uteri of implantation sites were examined by qRT-PCR. Interestingly, the expression levels of nesfatin-1/NUCB2 mRNA was significantly higher in abortion model mice than normal pregnant mice on day 14.5 of pregnancy (Fig. 4A). The levels of nesfatin-1/NUCB2 protein in the uteri of implantation sites were investigated by western blotting. Nesfatin-1/NUCB2 protein levels were increased in abortion model mice compared to normal pregnant mice, similar to nesfatin-1/NUCB2 mRNA expression (Fig. 4B).
DISCUSSION
Spontaneous abortion or miscarriage which is the most common complication of early pregnancy is defined as extraction of the embryo or fetus before 20-22 weeks of gestation (Baek et al., 2007). Spontaneous abortion is a multifactorial disorder resulting from genetic factors, anatomic factors, autoimmune disorders, endocrine dysfunction, thrombophilia, life style factors, and maternal infections (Wang et al., 2003; Ford & Schust, 2009). Furthermore, spontaneous abortion process involves a complex mechanism with various cytokines, growth factors, and hormones during the pregnancy (Feldt-Rasmussen & Mathiesen, 2011).
Nesfatin-1 is originally known for the first time to participate in the regulation of hunger as a neuropeptide produced in the hypothalamus of mammals (Oh-i et al., 2006). After that, many studies have shown that nesfatin-1/ NUCB2 is expressed not only in the hypothalamus, but also in the peripheral organs including gut, fat and heart (Mimee et al., 2012; Stengel et al., 2009a; Ramanjaneya et al., 2010). We also have reported that nesfatin-1/NUCB2 mRNA and protein is expressed in epithelial cells around the uterine glands and endometrium of mouse (Kim et al., 2014). Moreover, the nesfatin-1/NUCB2 expression was increased in the uterus of abortion model mice compared to those of normal pregnant mice (Chung et al., 2015). On the other hand, it has been reported that uterine nesfatin-1 expression levels are decreased significantly during pregnancy progresses in rat (Garces et al., 2014). These results suggest that nesfatin-1 protein in the uterus may play an important role during pregnancy and fetal development. However, little is known about how nesfatin-1/NUCB2 expressed in uterus is regulated. Therefore, we hypothesized that uterine nesfatin-1/NUCB2 expression could be regulated by cytokines in maternal serum during pregnancy. In this study, we examined the cytokine levels in maternal serum during pregnancy and the relationship between uterine nesfatin-1/NUCB2 expression and maternal serum cytokine levels using CBA/j × DBA/2 combination as a spontaneous abortion model mouse and CBA/j × BALB/c combination as a normal pregnant mouse.
We first investigated the number of implantation sites to confirm the pregnancy loss in the CBA/j × DBA/2 combination mice. Total number of implantation sites was significantly decreased in abortion model mice compared to normal pregnant. Next, we analyzed pro-and anti-inflammatory cytokine levels in the serum of normal pregnant and abortion model mice, and non-pregnant mice. IFN-γ, IL-1 and TNF are representative pro-inflammatory cytokines, and when they are administered to humans, they produce fever, inflammation, tissue destruction, and, in some cases, shock and death (Dinarello, 2000). Human pregnancy also increases the levels of IL-10, the anti-inflammatory cytokine that down regulates Th1 response as well as the release of pro-inflammatory cytokines, such as IFN-γ and TNF-α (Marzi et al., 1996; Jenkins et al., 2000). IFN-γ secreted in the uterus during early pregnancy plays critical roles that include initiation of endometrial vasculature remodeling, angiogenesis at implantation sites, and maintenance of the decidual (maternal) component of the placenta (Murphy et al., 2009). In our results, the pro-inflammatory cytokines, such as IFN-γ, IL-1α and TNF-α was significantly decreased in the serum of pregnant mice, and even in abortion model mice. However, their levels were not different between normal pregnant and abortion model mice. These results suggest that down-regulation of pro-inflammatory cytokines may be critical for the process of implantation and the maintenance of pregnancy, but they may be not related to spontaneous abortion.
On the other hand, our results showed that expression of anti-inflammatory cytokines, including IL-4, IL-13 and IL-1ra tended to be increased in normal pregnant mice, but their levels were significantly decreased in abortion model mice. The anti-inflammatory cytokines are a series of immunoregulatory molecules that control the pro-inflammatory cytokine response (Opal & DePalo., 2000). The production of pro-inflammatory cytokines coupled with a decrease or lack of increase in anti-inflammatory cytokines, such IL-4 and IL10 can lead to a spectrum of pregnancy disorders (Chatterjee et al., 2014). IL-13, predominantly secreted by activated Th2 cells, is also considered as an anti-inflammatory ‘IL-4-like’ molecule (Chomarat et al., 1998). Mononuclear lymphocytes, B cells, large granular lymphocytes and endothelial cells, known to be responsive to IL-13, can be traced in either the placenta or the maternal reproductive tract (McKenzie et al., 1993). IL-13 is thought to be required during the first third of gestation to support trophoblast invasion within the decidua (Zourbas et al., 2001).
In addition, we analyzed chemokines in the serum collected from normal pregnant and abortion model mice. Chemokines are a family of small cytokines having a role in leukocyte trafficking and participating in developmental processes such as differentiation and directed migration. Similar events occur in pregnancy during development of the fetal-maternal interface, where there is extensive leukocyte trafficking and tissue morphogenesis, and this is accompanied by abundant chemokine expression (Red-horse et al., 2004). We found that expression of SDF-1 and G-CSF was significantly decreased in both normal pregnant and abortion model mice, whereas M-CSF, IL-16, KC and MCP-1 were significantly decreased in abortion model mice. SDF-1/CXCR4 signaling promotes trophoblast survival during pregnancy, suggesting that alterations in SDF-1 and/or CXCR4 expression or function may be associated with specific pregnancy disorders (Jaleel et al., 2004). Granulocyte colony-stimulating factor (G-CSF), a cytokine, and its receptor are expressed in placental tissue. G-CSF treatment in women with spontaneous abortion showed to increase in the delivery of a healthy baby (Scarpellini & Sbracia, 2009). Macrophage colony stimulating factor (CSF-1 or M-CSF) is involved in haemopoiesis and probably in mouse gestation, showing that M-CSF increase early (4-8 weeks) and progressively during gestation. Sexual steroids induce its production by uterine glandular epithelial cells and its receptor is expressed on placental trophoblastic cells. This locally produced M-CSF could play a role in human pregnancy (Praloran et al., 1994). MCP-1 was higher in pregnant women (women with gestational diabetes mellitus and without) than in nonpregnant women. MCP-1 was elevated in patients with gestational diabetes mellitus (GDM) in the third trimester compared to healthy pregnant women, suggesting an association between inflammation and GDM (Klein et al., 2008).
In the present study, we also showed that nesfatin-1/NUCB2 mRNA and protein were expressed in uteri of implantation sites of normal pregnant and abortion model mice during the pregnancy. Interestingly, the expression levels of nesfatin-1/NUCB2 mRNA and protein were significantly higher in abortion model mice than normal pregnant mice. Given the cytokine array results, nesfatin-1/NUCB2 expression in the uterus of implantation site may be regulated cytokines and chemokines in the serum.
The present study demonstrated that serum levels of pro-inflammatory cytokines were not increased in abortion model mice, but anti-inflammatory cytokines were decreased compared to normal pregnant mice. In addition, serum chemokine levels also were decreased in abortion model mice compared to normal pregnant mice. By contrast, the expression levels of nesfatin-1/NUCB2 mRNA and protein in the uteri of implantation sites were significantly higher in abortion model mice than normal pregnant mice. These results suggest that uterine nesfatin-1/NUCB2 expression may be down-regulated by inflammatory cykokines (IL-4, IL-13, IL-1ra, IFN-γ, IL-1α and TNF-α) and chemokines (SDF-1, G-CSF, M-CSF, IL-16, KC and MCP-1) in the serum of pregnant mice. Moreover, this study suggests the possibility that nesfatin-1/NUCB2 expressed in the implantation sites may be associated with the maintenance of pregnancy.