
INTRODUCTION
Nesfatin-1 is a polypeptide encoded in the N-terminal region of the protein precursor, nucleobindin-2 (NUCB2), which also produces nesfatin-2 and -3 by the enzyme pro-hormone convertase (PC)-1/3 after putative post-translational processing (Miura et al., 1992; Barnikol-Watanabe et al., 1994). Nesfatin-1 has been known to have only physiological activity so far (Oh-I et al., 2006; Stengel et al., 2012; Schalla & Stengel, 2018).
Nesfatin-1 is initially known to be expressed in the hypothalamic nuclei such as arcuate nucleus, paraventricular nucleus, supraoptic nucleus, lateral hypothalamic area and zona incerta in rats (Brailoiu et al., 2007; Foo et al., 2008; Goebel et al., 2009; Xu et al., 2009; Stengel et al., 2010; Goebel-Stengel et al., 2011). Recent studies showed that a large amount of the nesfatin-1 expression was detected in the digestive system as well as in the brain. It has been also reported that the nesfatin-1 generated in the gastric organs, by acting in the brain, reduces food intake and suppresses the motilities of the stomach-duodenum (Jiang et al., 2016; Prinz et al., 2016). Nesfatin-1 immunoreactive cells are also co-localized with insulin in pancreatic beta-cells of mice and rats, suggesting a potential role for nesfatin-1 in pancreatic islet and glucose homeostasis (Gonzalez et al., 2009). Furthermore, nesfatin-1 was detected in the adipose tissues of humans and rodents, showing that it was expressed more in the subcutaneous than in the visceral fat (Shimizu et al., 2016).
Expression of NUCB2 mRNA has been detected in rat, mouse, dog, and human reproductive system (Garcia-Galiano et al., 2010; Garcia-Galiano et al., 2012; Nozawa et al., 2016). We also previously demonstrated that NUCB2/ nesfatin-1 is expressed in the mouse hypothalamus and pituitary and its expression levels were regulated by the gonadotropin (Kim et al., 2014; Chung et al., 2015, Seon et al., 2017). Moreover, the expression of NUCB2/nesfatin-1 in the pituitary and hypothalamus is regulated by estradiol and progesterone secreted from the ovary (Chung et al., 2015). Nesfatin-1 was also implicated in the maintenance of pregnancy because uterine expression levels of NUCB2/nesfatin-1 were increased in a mouse spontaneous abortion model (Chung et al., 2017).
Organogenesis is a crucial process during embryonic development of all animals, leading to the various organs in the body. Organogenesis starts with neural tube formation and cells in mesoderm layer are segmented into somites (Cunha & Baskin, 2016). The onset of mouse organogenesis has been known to begin at approximately E8.0, and the buds of all major organs are essentially formed at E9.5 (Cunha & Baskin, 2016). Around that time, many organogenesis-related genes and proteins are involved in the mouse fetal development and their interaction is crucial for the proper development of all organs (Baskin et al., 1996; Cunha & Baskin, 2016). Our previous reports have demonstrated that NUCB2/nestatin-1 was expressed in the fetus at embryonic day (E) 18.5 as well as neonatal and adult mice, suggesting that NUCB2/nestatin-1 may involved in the development and physiological function in mouse organs (Chung et al., 2013). Moreover, to further reinforce the hypothesis that NUCB2/nestatin-1 may involved in the organogenesis of mouse, it is needed to analyze expression in fetuses before E18.5 in mouse development. However, it is difficult to harvest organs of fetuses at E8.0 to E10.5, which is a critical time for mouse organogenesis, since the size of the fetus at that time is too small to obtain organs for qRT-PCR and Western blot analysis.
The aim of this study was therefore to investigate whether NUCB2/nestatin-1 is expressed in the fetuses earlier than E18.5, especially the fetus at E10.5. In this study, to identify the NUCB2 mRNA expression sites in specific organs, we focused on localization of NUCB2 mRNA in various organs of mouse fetus at E10.5 using in situ hybridization (ISH) method. In addition, we validated ISH results by immunofluorescence staining and qRT-PCR analysis.
MATERIALS AND METHODS
Pregnant ICR mice were purchased from Samtako Bio Korea (South Korea) and housed in groups of five per cage under controlled illumination (12:12 h light/dark cycle, lights on/off: 6 h/18 h) and temperature (22±2°). Animals were fed a standard rodent diet and tap water ad libitum. Animal care and experimental procedures were approved by the Institutional Animal care and the use committee at the Seoul Women's University in accordance with guidelines established by the Korea Food and Drug Administration.
To prepare sections of mouse fetus for ISH and IHC staining, pregnant mice at embryonic day (E)10.5 were euthanized by CO2 anesthesia. Fetuses were harvested after abdominal incision and fixed with 4% paraformaldehyde in HEPES buffer (30 mM HEPES, 100 mM NaCl, 2 mM CaCl2, pH 7.4). For paraffin embedding, fetuses were dehydrated in a graded series of alcohols, cleared with xylene, and embedded in paraffin (Paraplast Plus, Sigma, St. Louis, MO, USA). To prepare fresh organs of fetus at E13.5 and E17.5 for qRT-PCR, pregnant mice were euthanized by CO2 anesthesia and organs were quickly removed. The samples were kept in the ice and stored at −70° until using them.
The NUCB2 mRNA expression in the mouse fetus was examined by RNA ISH using the RNA scope technology (Advanced Cell Diagnostics, Hayward, CA, USA) according to the manufacturer's instruction. The paraffin embedded tissue section slides were baked for 1 h at 60° prior to use. After de-paraffinization and dehydration, the tissues were air dried and treated with hydrogen peroxide before boiling at 95° in a 1× Target Retrieval solution for 15 min. Protease was then applied for 30 min at 40°. NUCB2 probe was hybridized for 2 h at 40°, followed by a series of signal amplification which were hybridized to the target probes, and washing steps, and then color was developed with DAB. All hybridizations at 40° were performed in a HybEZ Hybridization System. Sections were then counterstained with 50% Mayer's hematoxylin diluted in distilled water for 2 min and dehydrated through ascending ethanol and xylene and Permanent Mounting Medium (Dako, CA, USA). Bright field images were acquired using a Leica microscope using a 40×objective. Negative control background staining was evaluated using a probe specific to the bacterial dapB gene.
The tissues embedded in paraffin were cut 10 μm sections using a microtome, deparaffinized, and rehydrated with graded xylene-alcohol series, and then washed with PBS before immunostaining. The sections were incubated with rabbit anti-rat nesfatin-1 polyclonal antibody (Phoenix Pharmaceuticals, INC., Burlingame, CA, USA) at 4° for overnight followed by incubation with Alexa fluor 594 conjugated goat anti-rabbit IgG (Jackson immuno research laboratory, West grove, PA, USA). The sections were counter stained with DAPI (4’,6-diamidino-2-phenylindole; Sigma, St. Louis, MO, USA) for 10 min and mounted on the slides with mounting medium (Vector laboratories, INC., Burlingame, CA, USA), and then observed under fluorescence microscopy (Axioskop2, Carl Zeiss, Germany).
Frozen organs were homogenized with 300 μL ice cold RNA isoplus (TaKaRa Bio, Shiga, Japan). After chloroform extraction and isopropyl alcohol precipitation, RNA was dissolved in 20 μL RNase-free DEPC (TaKaRa Bio, Shiga, Japan) solution. The RNA concentrations were measured with the Nano-drop (Thermo Fisher Scientific Inc., Waltham, MA, USA). First strand cDNA synthesis was performed using 2 μg RNA, 10 pmol oligo dT and RNase-free DEPC solution at 70° for 5min, followed by double-strand synthesis in 5× RT buffer (Invitrogen, Carlsbad, CA, USA) with 8 mM dNTP (BIO BASIC INC., Ontario, Canada), 200 unit/μL RTase (Invitrogen, Carlsbad, CA, USA) and RNase-free DEPC solution at 37° for 60 min and at 72° for 15 min. qRT-PCR was performed in a total volume of 20 μL buffer solution containing 2 μL of template cDNA, 10 μL of SYBR Green (Roche, Manheim, Germany), and 10 pmol of each primer. Primer pairs were as follows: NUCB2 forward 5’-AAAACCTTGGCCTGTCTGAA-3’; reverse 5’-CATCGATAGGAACAGCTTCCA-3’ and GAPDH forward 5’-TTGATGGCAACAATCTCCAC-3’; reverse 5’-CGTCCCGTAGACAAAATGGT-3’ (BIONICS, Seoul, Korea). The optimal temperature cycling protocol was determined to be 95° for 5 min followed by 45 reaction cycles at 95° for 10 s, 60° for 10 s and 72° for 10 s using the LightCycler® 480 Real-time PCR System (Roche, Manheim, Germany).
RESULTS
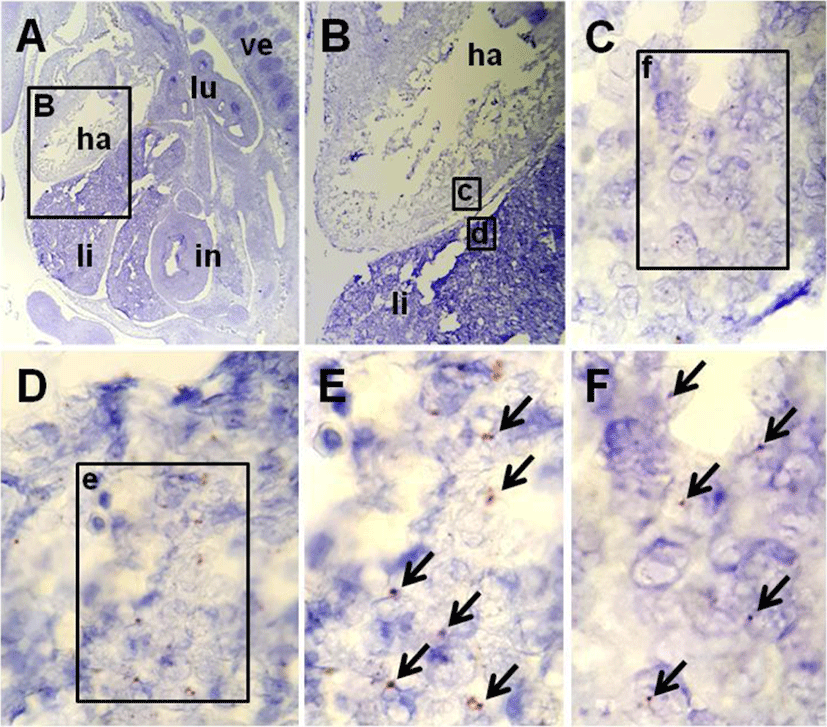
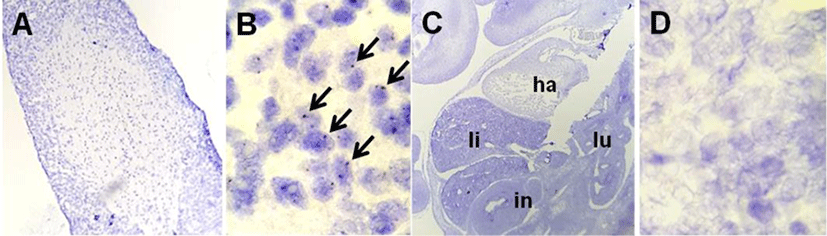
The localization of NUCB2 mRNA in the mouse fetus was verified by a highly-sensitive ISH system using NUCB2 probe. The positive signals were present as punctate cytoplasmic and/or nuclear brown staining that was higher than the signals on the negative control slide (Fig. 2C, D). Fig. 1 shows that NUCB2 mRNA was localized in the liver and heart of mouse fetus at E10.5. Many of dark brown dots as positive signal were observed in the liver (Fig. 1D, E), while relatively less positive signals were in the heart (Fig. 1C, F). On the other hand, no positive signals were detected in other organs of the mouse fetus at E10.5. As a positive control for NUCB2 mRNA ISH, we used the pituitary gland of adult mouse, in which NUCB2/nesfatin-1 is expressed a lot (Kim et al., 2014; Chung et al., 2015). Strong positive signals in pituitary gland slides established the validity of ISH results in the liver and heart (Fig. 2A, B). The pituitary gland of adult mouse was used as a positive control for ISH.
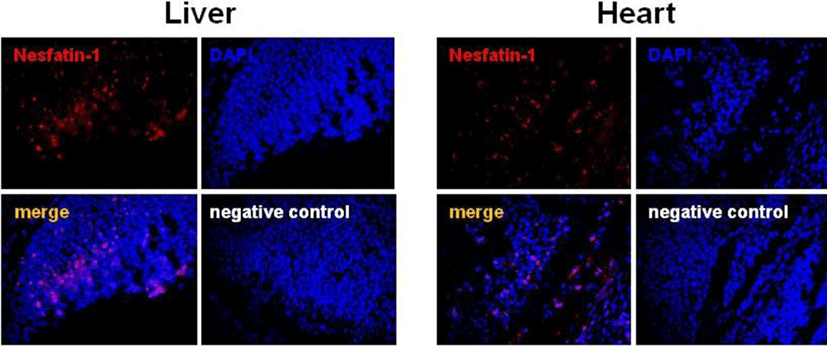
The localization of NUCB2 mRNA in the mouse fetus was further verified by immunofluorescence staining for nesfatin-1 protein. In accordance with ISH results, nesfatin-1 protein was expressed in the liver and heart of the mouse fetus at E10.5, where NUCB2 mRNA was detected by ISH (Fig. 3). Many cells stained with red fluorescence were observed in the liver and heart, whereas no cells stained with red were detected on the negative control slide (Fig. 3).
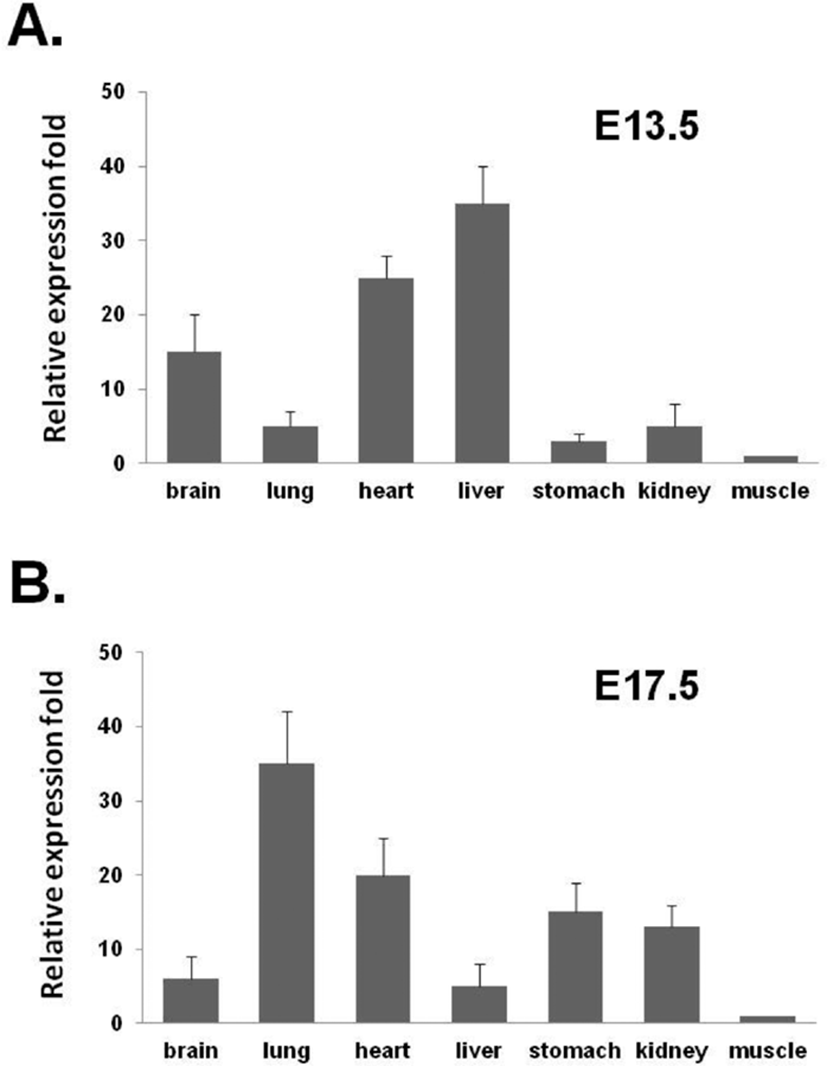
The expression of NUCB2 mRNA in the various organs of the mouse fetus at E13.5 and E17.5 were measured by qRT-PCR and compared the differences between the two fetuses. In the mouse fetus at E13.5, the levels of NUCB2 mRNA were relatively higher in liver and heart than in other organs, but low in lung, stomach, and kidney (Fig. 4A). On the other hand, in the mouse fetus at E17.5, the levels of NUCB2 mRNA were relatively higher in lung, heart, stomach, and kidney than in other organs, but low in liver (Fig. 4B). Interest ingly, NUCB2 mRNA expression in the liver was dramatically reduced in the fetus at E17.5 compared to the fetus at E13.5, whereas its expression levels in lung, stomach, and kidney were increased in the fetus at E17.5 (Fig. 4).
DISCUSSION
We previously reported that NUCB2/nestatin-1 is expressed in the various organs of fetuses and neonates as well as adult mice. In adult mice, NUCB2 mRNA was mainly expressed in the pituitary and at relatively lower levels in the hypothalamus, spleen, thymus, heart, liver, and muscle (Chung et al., 2013; Kim et al., 2014). Interestingly, expression in reproductive organs such as the testis, epididymis, ovary, and uterus was significantly higher than in the hypothalamus (Chung et al., 2013; Kim et al., 2014). On the other hand, NUCB2/nesfatin-1 expressed in the fetus and neonate was relatively higher in lung, kidney, and stomach than other organs. Comparing the level of NUCB2 mRNA expression in fetuses and neonates with that in adult mice, expression level was decreased in the stomach, but increased in the kidney of adult mice. These results suggest that NUCB2/ nesfatin-1 may be involved in the development and physiological function of mouse organs (Chung et al., 2013).
In previous study, we used the fetus at embryonic day (E) 18.5 to examine the levels of NUCB2 mRNA and nesfatin-1 protein expression in the organs (Chung et al., 2013). It is well known that the formation of main organs during mouse fetal development begins at E8.0 to E10.0 (Tam & Behringer, 1997; Zorn & Wells, 2009). Considering this fact, it is critical to examine the expression of organogenesis-related genes during this period when organ formation begins. However, the size of the fetus at about E10.0 when organ formation begins is too small to obtain organs. Thus, it is impossible to examine gene expression by qRT-PCR analysis. To overcome this obstacle, we used high-sensitive ISH technique instead of qRT-PCR. The advantage of ISH is that it can identify specific gene expression sites where it is expressed at even low levels. Moreover, it is not necessary to harvest fetal organs, since tissue sections are used in ISH. (Baumgart et al., 1997; Chotteau-Lelievre et al., 2006; Stylianopoulou et al., 2012). Considering these advantages of ISH, we performed ISH to examine NUCB2 mRNA expression in the fetal organs at E10.5. As a result of ISH, NUCB2 mRNA was localized in the liver and heart of mouse fetus at E10.5, whereas no positive signals were detected in other organs. To further verify the localization of NUCB2 mRNA expression in the liver and heart, we next performed immunofluorescence staining for nesfatin-1 protein encoded in NUCB2 gene. In accordance with ISH results, nesfatin-1 protein was expressed in the fetal liver and heart. The fact that NUCB2 mRNA and nesfatin-1 protein were expressed in the same sites indicates that NUCB2 mRNA is certainly expressed in the liver and heart. Given the ISH and IHC results, NUCB2/nesfatin-1 may be involved in the development of the liver and heart at the beginning of organ formation.
Similar to our study, gene expression of leptin known to involve in the regulation of energy balance and appetite like nesfatin-1 was localized to specific sites in the mouse fetus by ISH. High levels of gene expression of leptin were observed in the placenta, fetal cartilage, and hair follicles in the mouse fetus at E14.5, suggesting that leptin may play a role in the growth and development of the fetus (Hoggard et al., 1997; Warchol et al., 2014). Ghrelin is also a participant in regulating the complex process of energy homeostasis by showing the opposite function of nesfatin-1 to appetite. At early stages of mouse fetal development, ghrelin was expressed in the primitive epithelium of lungs, and decreased in levels of expression with gestational age. The fetal lung is one of the first to express ghrelin during development and might be considered a new source of circulating fetal ghrelin (Santos et al., 2006; Warchol et al., 2014). Taken together, these results suggest nesfatin-1 may also be involved in the growth and development of mouse fetus.
To determine the level of NUCB2 mRNA in the fetuses as early as possible to get organs at the developmental stage of the moue, we used the fetuses at E13.5. In addition, their levels of NUCB2 mRNA were compared with those of the fetuses at E17.5. In the mouse fetus at E13.5, the levels of NUCB2 mRNA were relatively higher in liver and heart than in other organs, but low in lung, stomach, and kidney similar to ISH result. On the other hand, in the mouse fetus at E17.5, the levels of NUCB2 mRNA were relatively higher in lung, heart, stomach, and kidney than in other organs, but low in liver. In our previous study using the mouse fetus at E18.5, the levels of NUCB2 mRNA and nesfatin-1 protein expressed in the organs were also relatively high in lung, heart, stomach, and kidney but low in liver, similar to those of in mouse fetus at E17.5 (Chung et al., 2013). These differences in the expression of NUCB2/nesfatin-1 between the two fetuses at the developmental stage suggest that NUCB2/ nesfatin-1 may play an important role in organogenesis during mouse fetal development.
We demonstrated here for the first time that NUCB2 mRNA was localized in the liver and heart of mouse fetus at E10.5 by ISH and nesfatin-1 protein was detected in the same organs by IHC. In addition, the levels of NUCB2 mRNA expressed in liver and heart at E13.5 were different compared to that of the mouse fetus at E17.5. These results suggest that NUCB2/nesfatin-1 may play an important role in the development and physiological function of liver and heart during organogenesis in the mouse fetus. In the future, we need more study on the function of NUCB2/nesfatin-1, which is highly expressed in the liver and heart at the beginning of organogenesis during mouse fetal development.