INTRODUCTION
In 2020, aquaculture in Korea yielded 325,889 tons of oysters (Korean Statistical Information Service, Ministry of Oceans and Fisheries, 2020). The main production site is the south coast, while some varieties are cultivated via various methods on the west coast (Hwang et al., 2016). However, the seedlings needed for oyster farming in Korea are dependent on natural culture. When there are few natural seedlings, it is difficult to secure a stable seed supply. Since the 1970s, the pollution of coastal waters, aging of fish farms, and deterioration of fishing conditions have led to seed shortages. In 1992, a major seed-collecting area on the southern coast was severely damaged (Park et al., 1999). Currently, artificial seed production is actively conducted to ensure a stable supply to meet the demand for oysters. The quality of the mother oysters affects seedling growth (Lee et al., 2008; Kim et al., 2013). Although studies have evaluated mother oysters by examining changes in body components (Lim et al., 2011; Kim et al., 2014), a few studies have evaluated endogenous factors, such as the expression of insulin-like growth factors (IGFs), in mother oysters. The IGF system regulates somatic growth and development in fish, as in mammals (Duan, 1997; Le Bail et al., 1998; Moriyama et al., 2000). IGF-1 is an anabolic hormone that regulates development, somatic and skeletal muscle growth, and reproduction via endocrine, autocrine, and paracrine mechanisms mediated by growth hormone (Duan, 1997; Liu et al., 2017).
Here, we examined IGF-1 receptor (IGF-1R) expression and the phosphorylation of endogenous substrates in the IGF-1 pathway to evaluate the growth and maturity of artificially matured female and male Pacific oysters.
MATERIALS AND METHODS
Broodstock of artificially matured Pacific oysters, C. gigas, was obtained from the Southeast Sea Fisheries Institute, National Institute of Fisheries Science, Namhae, Korea. Each month from March to June 2016, 80 C. gigas were obtained. These were reared in running water at a flow rate of 15 L/min and fed a mixture of 10–15×104 cells/mL Isochrysis galbana, Pheodactylum sp., Tetraselmis sp., and Chlorella sp. The water temperature was raised slowly from 14.5℃ to 22.5℃ over 15 days and then kept at 22.5±0.34℃ until the end of the study. The salinity was 32.2–33.8 psu.
The oyster shell length (SL), shell height (SH), and shell width (SW) were measured using Vernier calipers; total wet weight and soft tissue weight (STW) were measured using an electronic balance. The condition index (CI) and tissue weight rate (TWR) were calculated according to Choi & Chang (2003) using the following equations:
The adductor muscle, digestive gland, gills, labial palps, mantle edge, and gonads were isolated, and 0.5 g of each was frozen in liquid nitrogen and stored at −80℃.
The tissues were homogenized in RIPA lysis buffer (50 mM Tris, 1 mM EGTA, 150 mM NaCl, 1% NP-40, and 0.25% sodium deoxycholate) containing protease inhibitors (1 μg/mL aprotinin, 1 μg/mL leupeptin, 1 μg/mL pepstatin, 1 mM Na3VO4, 1 mM NaF, and 1 mM PMSF). Samples were centrifuged at 14,483×g for 10 min, and the supernatant was collected. The protein concentration was measured using a BCA protein assay kit (Pierce Biotechnology, Rockford, IL, USA). Proteins (35–50 μg) were separated on 8%–10% polyacrylamide gels by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to polyvinyl difluoride membranes (Millipore). The membranes were blocked in TBS-T (10 mM Tris-HCl, 150 mM NaCl pH 7.5, and 0.1% Tween 20) containing 1% BSA, incubated with the primary and secondary antibodies at room temperature for at least 1 h, and then detected using enhanced chemiluminescence Western blotting reagents (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Between each step, the membranes were washed twice with TBST. The following primary antibodies and an anti-mouse secondary antibody, obtained from Santa Cruz Biotechnology, were diluted 1:1,000: IGF-1, IGF-1R, phosphorylated (p)-IGF-1R (PY99), AKT, p-AKT, ERK, and p-ERK. Monoclonal mouse anti-β-actin antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) was used as a control. The proteins were detected using enhanced chemiluminescence detection reagents (Advansta, Menlo Park, CA, USA) and visualized using the GeneSys imaging system (Syngene Synoptics, London, UK).
RESULTS AND DISCUSSION
The growth of C. gigas was assessed by measuring SL, SH, SW, and STW for 4 months (Fig. 1A). Beginning in April, when it is possible to distinguish males and females, the monthly CI and TWR were assessed (Fig. 1B). SL, SH, SW, and STW were all significantly (p<0.05) lower in March when maturation induction was started. Shell growth did not differ until June, but STW increased over time (Fig. 1A). In March, it was difficult to distinguish male and female oysters. While there was no difference in shell growth from April to June, females and males had the highest CI and TWR in June, when the water temperature had been elevated the longest (Fig. 1B).
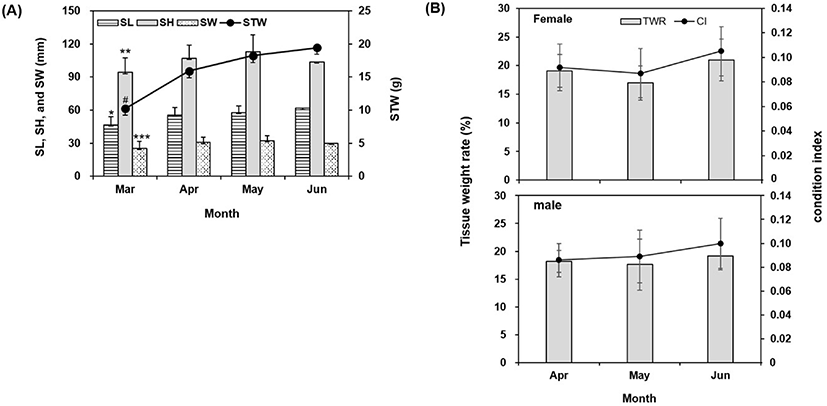
Oyster maturation depends on water temperature, which affects gamete maturation, ovulation, and spawning in many fish species and oysters (Loosanoff & Davis, 1952; Anguis & Canavate, 2005; De Oliveira Ramos et al., 2014). Oyster maturation can depend on temperature, even within the genus Crassostrea (Loosanoff & Davis, 1952; De Oliveira Ramos et al., 2014). Mangrove oyster C. gasar gonadal tissue matured at 22℃ and 26℃, but not at 18℃ (De Oliveira Ramos et al., 2014), and the CI did not differ among the temperature treatments at the beginning of the experiment.
In oysters of the genus Crassostrea, an individual can be male or female depending on the environmental conditions (De Oliveira Ramos et al., 2014). In this study, the mean ratio of C. gigas females to males from April to June was 1:0.875.
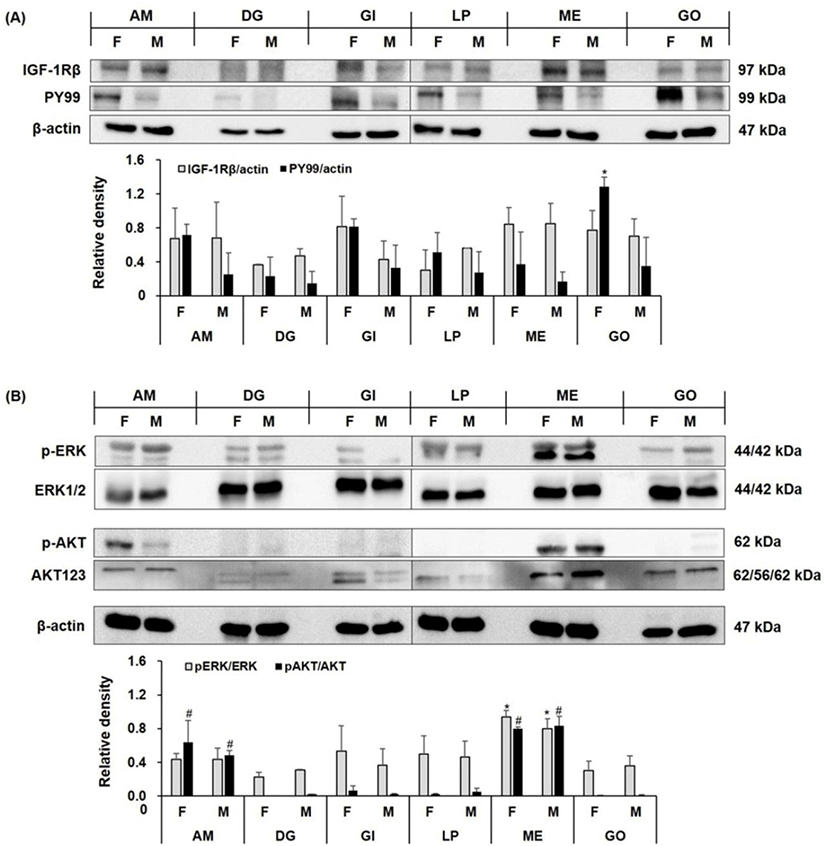
IFG-1R has two extracellular α-subunits with ligand-binding sites. Each α-subunit couples to one of two membrane-spanning β-subunits, which contain an intracellular domain with intrinsic tyrosine kinase activity (Adams et al., 2000). IGF-1 binding to its receptor initiates a complex signaling cascade in the cell (Foncea et al., 1997). Activation is initiated by triggering the β-subunit kinase domains, leading to receptor autophosphorylation and tyrosine phosphorylation of multiple substrates (Adams et al., 2000). IGF-1R activation may be involved in the survival of surf clam (Spisula sachalinensis) larvae (Choi & Nam, 2014) and the growth of C. gigas (Choi et al., 2018). We investigated the expression of IGF-1R and the phosphorylation of its endogenous substrates in various tissues, including the gonads of oysters artificially matured via water temperature elevation. IGF-1Rβ was detected in all female and male tissues, whereas tyrosine phosphorylation (PY99) of the intracellular β-subunits was clearly detected in females, especially the gonads (p<0.05, Fig. 2A). There was significant p-ERK and p-AKT expression in the adductor muscle and mantle edge (p<0.05, Fig. 2B). While there were differences in the expression of endogenous substrates, the expression of downstream factors is believed to be involved in IGF signaling in artificially matured oysters. Further studies are needed to examine the association between the IGF-1 signaling pathway (PI3K and Ras/MAPK pathways) and maturation mechanisms in shellfish.