INTRODUCTION
The Drosophila testis is a powerful model for investigating the establishment, maintenance, and age-related deterioration of the stem cell niche and its regulation of resident stem cells (Matunis et al., 2012; Sinden et al., 2012; Voog et al., 2014; Anllo et al., 2019; Kong et al., 2024). At the apical tip of the testis, a cluster of approximately 10 hub cells forms a specialized structure that provides a niche for two stem cell populations: germline stem cells (GSCs) and cyst stem cells (CySCs) (Fuller & Spradling, 2007; Voog et al., 2014; Greenspan et al., 2015). Hub cells support stem cell maintenance through direct, adhesion-mediated contact and by secreting niche-derived signaling molecules, most notably Unpaired (Upd), which activates Janus kinase/signal transducer and activator of transcription (JAK/STAT) signaling to promote stem cell self-renewal (Kiger et al., 2001; Tulina & Matunis, 2001; Yamashita et al., 2005; Leatherman & Dinardo, 2008; Inaba et al., 2010). Physical association with the hub orients the asymmetric divisions of both GSCs and CySCs; in each lineage, the daughter cell that remains attached to the hub retains stem cell identity, whereas the displaced daughter initiates differentiation (Yamashita & Fuller, 2008; Inaba et al., 2010; de Cuevas & Matunis, 2011). GSC daughters become gonialblasts (GBs), which undergo four rounds of transit-amplifying mitoses to generate 16-cell spermatogonial cysts that subsequently differentiate into spermatocytes and enter meiosis to produce 64 haploid sperm. Throughout germline development, cyst cells derived from CySCs encapsulate and support the germline cyst, coordinating differentiation and morphogenesis (Fuller & Spradling, 2007; Matunis et al., 2012).
Recent studies have identified the evolutionarily conserved RNA-binding proteins Lin28 and IGF-II mRNA-binding protein (IMP) as key intrinsic regulators of hub cell maintenance and niche function (Toledano et al., 2012; Sreejith et al., 2019; To et al., 2021; Sreejith & Kim, 2023). Lin28 is best known for inhibiting let-7 microRNA biogenesis by promoting the uridylation and degradation of let-7 precursors, thereby protecting let-7 target genes from silencing (Heo et al., 2008; Viswanathan et al., 2008; Heo et al., 2009; Yamashita et al., 2019; Shaik Syed Ali et al., 2025). In hub cells, Lin28 levels are high in young males and decline with age (Sreejith et al., 2019), whereas let-7 expression exhibits the inverse trend, increasing in older hubs (Toledano et al., 2012), suggesting that Lin28 regulates let-7 in hub cells. To test the hypothesis that Lin28 represses multiple microRNAs, including let-7, in hub cells, we employed microRNA expression profiling of wild-type and lin28 mutant testes. MicroRNA array analysis revealed 20 microRNAs upregulated in lin28 mutants, including let-7, demonstrating that the low levels of let-7 in young hub cells are due to repression by Lin28. Here, we report genetic analysis of miR-304, which was identified through this profiling.
MATERIALS AND METHODS
Flies were reared on standard cornmeal/agar medium supplemented with yeast at 25°C, 60±5% relative humidity, under a 12 h light/12 h dark cycle. The following stocks were obtained from the Bloomington Drosophila Stock Center: w1118, lin28EP915 (BDSC #17298), and miR-304 knockout mutant (BDSC #58918).
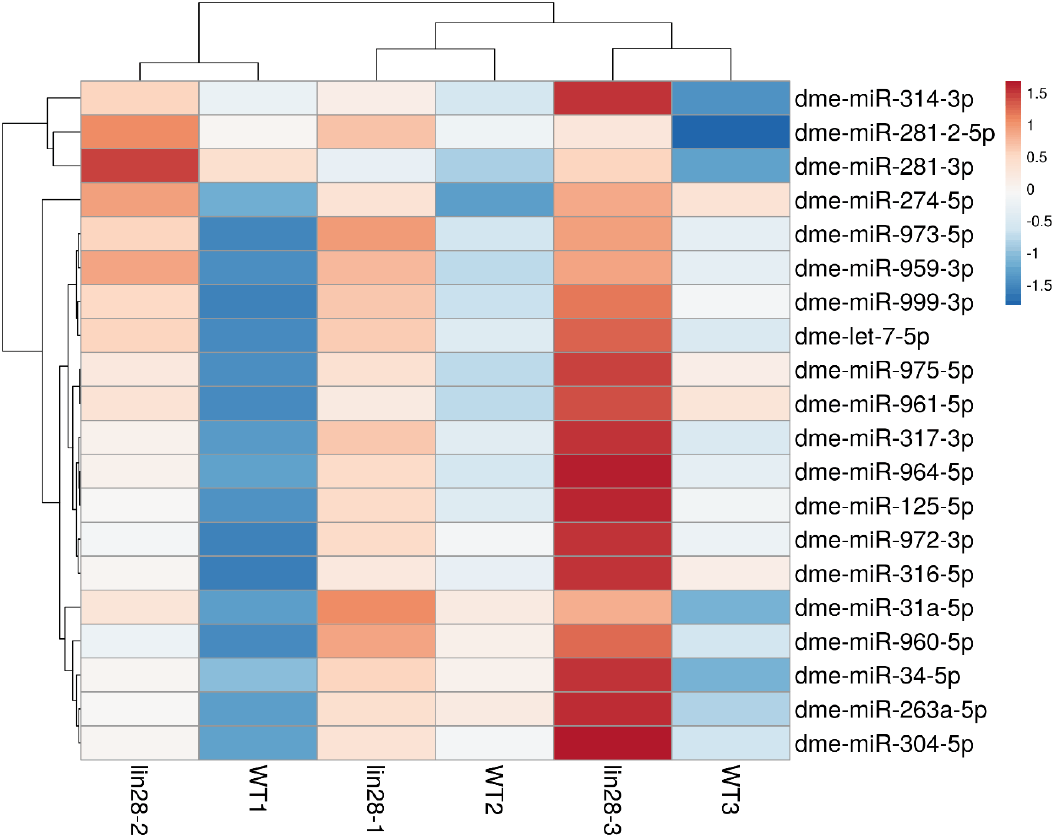
Testes from 0–1-day-old males (w1118 and lin28EP915) were dissected in sterile, ice-cold 1× phosphate-buffered saline (PBS) in triplicate. Dissected testes were immediately flash-frozen in liquid nitrogen prior to RNA extraction. Briefly, RNA extraction was performed in two steps. First, testes were lysed in lysis buffer followed by phenol:chloroform extraction. RNA was then further purified using the Ambion microRNA extraction kit. Purified RNA was shipped to Exiqon Services (Exiqon, Vedbæk, Denmark) on dry ice for microRNA profiling. All microRNA array experiments were conducted at Exiqon Services. For each sample, 750 ng total RNA (sample and reference) was labeled with Hy3™ and Hy5™ fluorescent dyes, respectively, using the miRCURY LNA™ microRNA Hi-Power Labeling Kit, Hy3™/Hy5™ (Exiqon), following the manufacturer’s instructions. Hy3™-labeled samples and Hy5™-labeled reference RNA were mixed pairwise and hybridized to the miRCURY LNA™ microRNA Array (Drosophila melanogaster) (Exiqon), which contains capture probes targeting microRNAs registered in miRBase v18.0. Hybridization was performed according to the miRCURY LNA™ microRNA Array instruction manual using a Tecan HS4800™ hybridization station (Tecan, Männedorf, Austria). After hybridization, microarray slides were scanned and stored in an ozone-free environment (ozone level<2.0 ppb) to prevent bleaching of fluorescent dyes. Slides were scanned using the Agilent G2565BA Microarray Scanner System (Agilent Technologies, Santa Clara, CA, USA), and image analysis was performed using ImaGene® 9 (miRCURY LNA™ microRNA Array Analysis Software, Exiqon). Quantified signals were background-corrected using Normexp with an offset of 10 and normalized using global locally weighted scatterplot smoothing (LOWESS) regression. Upregulated microRNAs in lin28 mutant testis relative to wild-type testis were visualized in a heatmap, with values representing fold change. Specifically, values of 0.5, 1, and 1.5 correspond to 1.4-, 2-, and 2.8-fold increases, respectively. The microRNA data have been deposited in ArrayExpress (E-MTAB-17143, Hinxton, UK).
Testes from 2~3-day-old males were dissected in 1× PBS. The following primary antibodies were used: mouse monoclonal anti-Fasciclin III [FasIII; 1:50 dilution; Developmental Studies Hybridoma Bank (DSHB, Iowa City, IA, USA) 7G10], rabbit polyclonal anti-IMP (1:500 dilution; a gift from Dr. Paul Lasko), and guinea pig polyclonal anti-Traffic Jam (TJ; 1:2,000 dilution; a gift from Dr. Dorothea Godt). Following primary antibody incubation, samples were washed twice for 15 min in phosphate-buffered saline with tween (PBST) and incubated with secondary antibodies in blocking solution for 2 h at room temperature. Secondary antibodies (all from Invitrogen, unless otherwise noted) included: Alexa Fluor 488-conjugated goat anti-rabbit (A11008, 1:800); Alexa Fluor 488-conjugated goat anti-mouse (A11001, 1:800); Alexa Fluor 555-conjugated donkey anti-mouse (A31570, 1:800); Alexa Fluor 555-conjugated goat anti-rabbit (A21429, 1:800); and Cy3-conjugated goat anti-horseradish peroxidase (HRP) (Jackson ImmunoResearch, West Grove, PA, USA, 1:800). Nuclei were stained with 4’,6-diamidino-2-phenylindole (DAPI; 1:1,000) for 30 min. Finally, samples were washed twice in PBST and mounted using Fluoromount-G (SouthernBiotech, Birmingham, AL, USA). Confocal images were acquired using a Leica Application Suite X (LAS X, Leica Microsystems GmbH, Wetzlar, Germany) confocal microscope. Image processing and analysis were performed using Leica LAS X and ImageJ/Fiji software (National Institutes of Health, Bethesda, MD, USA). To quantify fluorescence intensity in the hub cells, the hub area was outlined using ImageJ, and the mean fluorescence intensity was measured. To account for variations in sample preparation, these values were normalized to the background signal measured in non-expressing regions within the same sample.
Putative microRNA binding sites in the 3’ untranslated regions (3’ UTRs) of candidate target mRNAs were predicted using TargetScan Fly (version 7.2; http://www.targetscan.org/fly/, Whitehead Institute for Biomedical Research, Cambridge, MA, USA) and miRanda (version 3.3a; http://www.microrna.org, Memorial Sloan Kettering Cancer Center, New York, NY, USA). Predictions were based on seed sequence complementarity (nucleotides 2–7 or 2–8 of the mature microRNA).
Statistical analyses were performed using GraphPad Prism 11.0.2 software (GraphPad Software, LLC, Boston, MA, USA). Data are presented as the mean±SEM from three independent testes (n=3). Differences between two groups were determined using a two-tailed unpaired Student’s t-test. Statistical significance is denoted as follows: ns, p>0.05; **** p<0.0001.
RESULTS
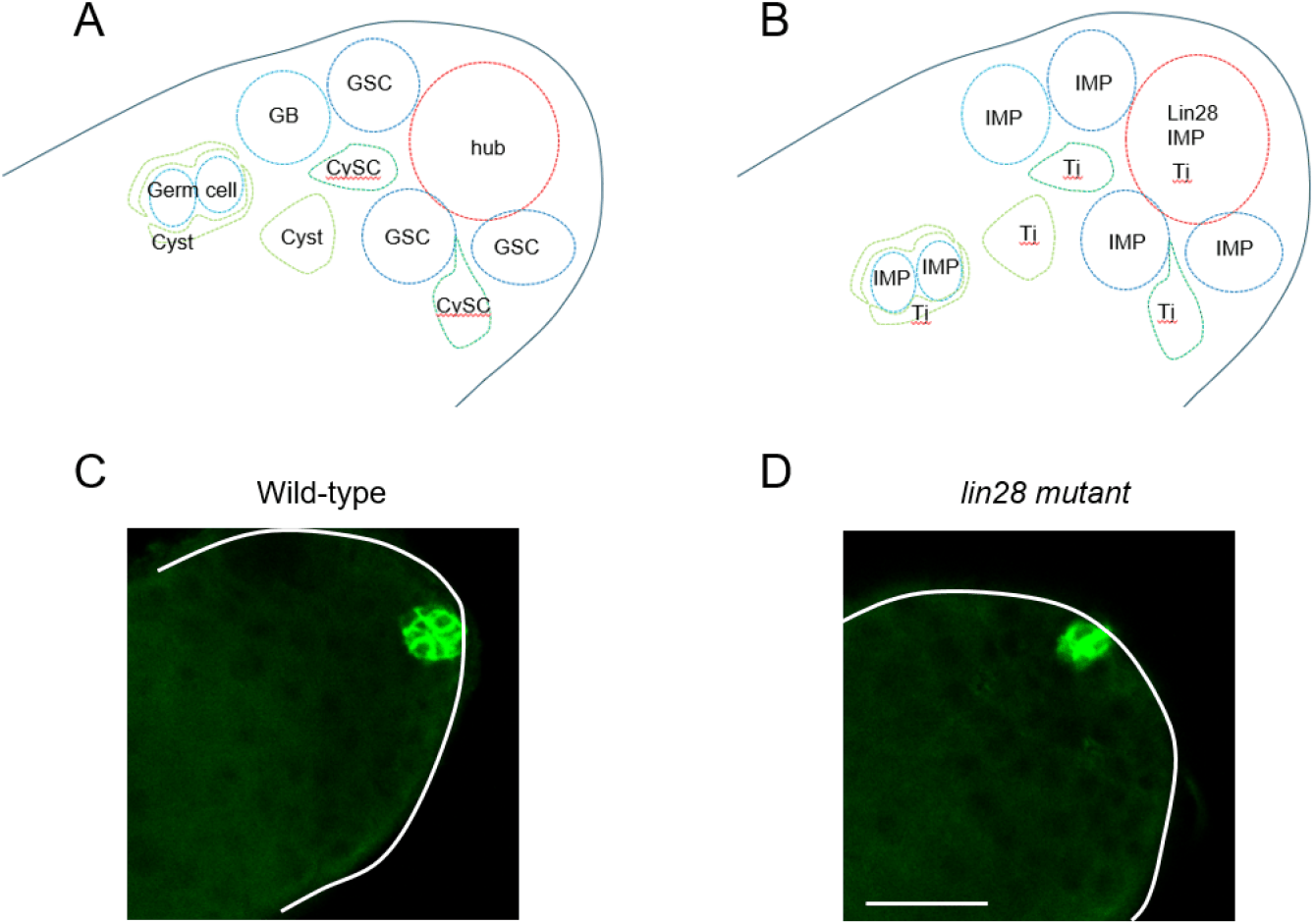
The apical tip of the Drosophila testis comprises diverse somatic cells, including hub cells, CySCs, and cyst cells, alongside germline cells, including GSCs and GBs (Fig. 1A) (Matunis et al., 2012). Lin28 is exclusively expressed in hub cells and is absent in other testicular cell types (Fig. 1B) (Sreejith et al., 2019). As previously reported, lin28 mutant testes exhibit hub cell loss, a defective hub structure (Fig. 1C and D) (Sreejith et al., 2019), suggesting that Lin28 plays a crucial role in maintaining hub identity and niche function. Given that the best-characterized biochemical role of Lin28 is to mediate the degradation of the microRNA precursor pre-let-7, we hypothesized that the hub defects in lin28 mutant testes might be caused by the upregulation of let-7 and other microRNAs. To test this, we performed microRNA array profiling of wild-type and lin28 mutant testes. The resulting heatmap identified 20 microRNAs that were upregulated in lin28 mutant testes, including let-7 (Fig. 2), demonstrating that Lin28 represses let-7 and multiple other microRNAs in young hub cells.
It was previously shown that let-7 targets the mRNA of the intrinsic hub factor imp (Toledano et al., 2012). The IMP protein protects upd mRNAs from degradation by siRNAs in hub cells (Toledano et al., 2012). Upd, a hub-specific self-renewal signaling molecule, is secreted to stimulate the JAK/STAT pathway in adjacent stem cells, which is required for stem cell self-renewal (Kiger et al., 2001; Tulina & Matunis, 2001; Boyle et al., 2007; Issigonis et al., 2009). Thus, our microRNA profiling, which identified let-7 as upregulated in lin28 mutant testes, suggests that Lin28 represses let-7 expression in hub cells to protect imp mRNAs from let-7-mediated repression.
Among the microRNAs upregulated in lin28 mutant testes, we selected miR-304 for further analysis. First, miR-304 contains a predicted seed site in the 3› UTR of imp (Fig. 2) (Toledano et al., 2012). Second, a miR-304 mutant was available from the Drosophila stock center. Third, IMP expression can be monitored using an anti-IMP antibody. These factors enabled us to examine whether miR-304 targets imp. We immunostained wild-type and miR-304 mutant testes with antibodies to identify specific cell types. In particular, anti-FasIII antibody marks hub cells (Fig. 1C and D).
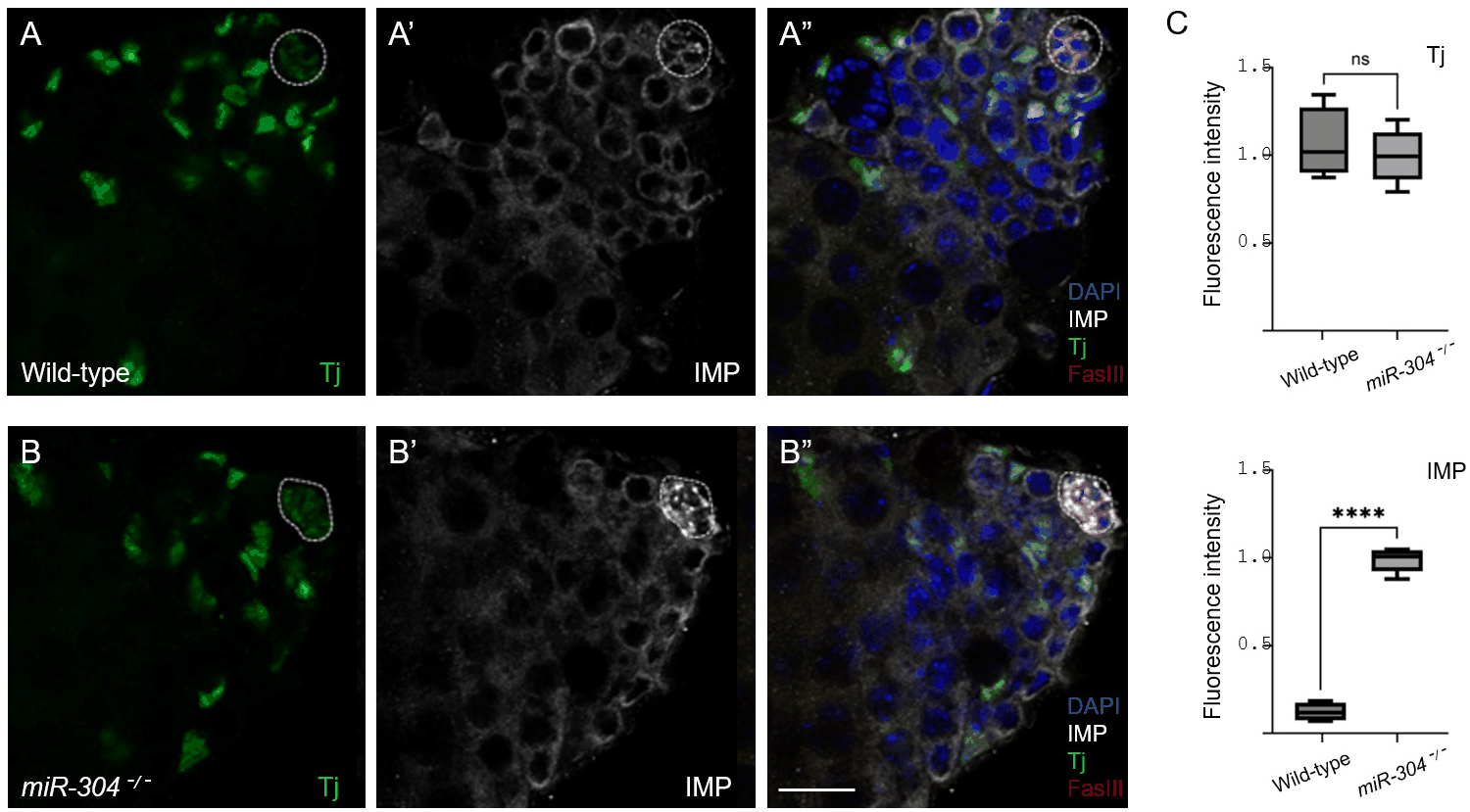
Immunohistochemical staining revealed that IMP protein levels as visualized by immunofluorescence intensity were noticeably increased specifically in the hub cells of miR-304 mutant testes compared to wild-type controls (Fig. 3A’, B’, and C). The IMP signal intensity in germ cells appeared unchanged (Fig. 3A’ and B), demonstrating that miR-304 exerts a hub-restricted effect on imp mRNA, which is consistent with the hub-specific expression of Lin28 (Sreejith et al., 2019). As a control, we stained testes with an anti-Traffic jam (Tj) antibody, which labels Tj-expressing cells, including hub cells and various somatic cells (Li et al., 2003; Wingert & DiNardo, 2015). We observed no significant difference in Tj staining intensity in Tj-positive somatic cells between wild-type and miR-304 mutants (Fig. 3A–C), highlighting the specificity of miR-304 in restricting IMP, but not Tj, expression in hub cells. Thus, our microRNA profiling, which identified miR-304 as upregulated in lin28 mutant testes, suggests that Lin28 represses miR-304 expression in hub cells to protect imp from miR-304-mediated repression.
DISCUSSION
Our microRNA array analysis revealed that lin28 mutant testes contain elevated levels of multiple microRNAs compared to wild-type controls. Given that Lin28 expression in the testis is restricted to hub cells (Sreejith et al., 2019), this upregulation likely reflects the loss of Lin28-mediated repression of these microRNAs within the niche.
Our profiling identified let-7 as upregulated in lin28 mutant testes, suggesting that Lin28 represses let-7 expression in hub cells. These findings are consistent with the known expression dynamics of Lin28 and let-7: Lin28 is highly expressed in the young testis and decreases gradually, whereas let-7 shows an inverse expression pattern (Toledano et al., 2012; Sreejith et al., 2019). Thus, the accumulation of let-7 in aged testes may, in part, result from the age-dependent decline of Lin28 (Sreejith et al., 2019). Since let-7 is known to target imp mRNA, this regulatory pathway accounts for the decreased IMP levels observed in aged testes (Toledano et al., 2012). Here, we demonstrate that miR-304, another microRNA identified as upregulated in lin28 mutant testes, also targets imp mRNA. This conclusion is supported by miR-304 mutant analysis; however, future research utilizing UAS-miR-304 transgenic lines and reporter constructs (GFP) reporter constructs containing miR-304 target sequences will be required to fully establish this relationship.
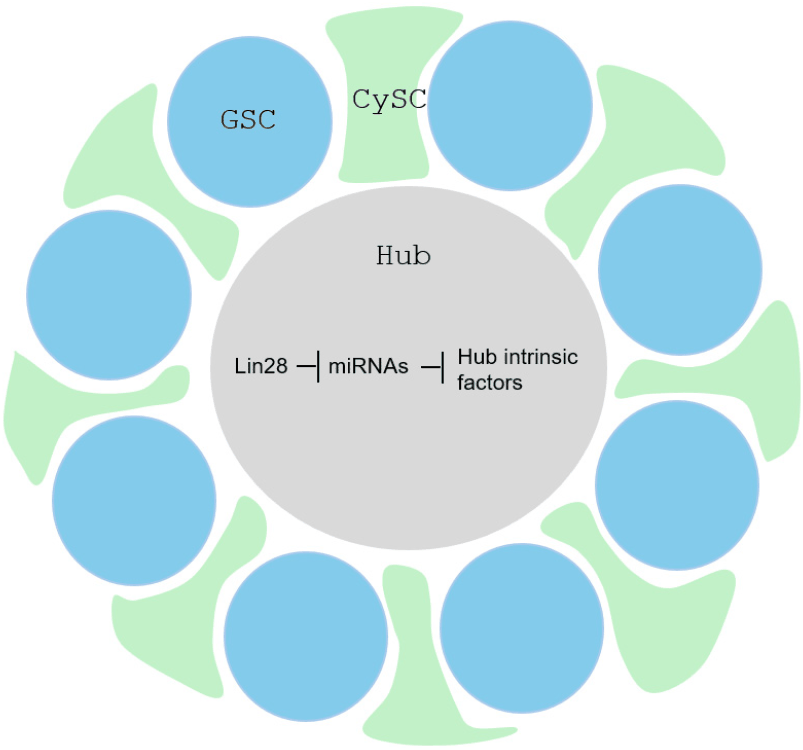
IMP has recently been identified as a critical intrinsic hub factor required for the stability of upd transcripts, which encode a key self-renewal signal expressed in hub cells (Toledano et al., 2012). Thus, our study suggests that Lin28 maintains IMP and Upd expression in hub cells through the coordinated repression of let-7 and miR-304. Additionally, our profiling identified a suite of other microRNAs upregulated in lin28 mutant testes, whose roles remain unexplored. We propose that Lin28 represses these microRNAs to protect other, as-yet-unidentified essential hub factors (Fig. 4). Future research will focus on characterizing these additional microRNA targets to further elucidate the gene regulatory networks through which Lin28 maintains the stem cell niche.