
INTRODUCTION
Reproductive processes and early embryonic development are maintained by sophisticated cell signaling, hormonal regulation, and intercellular interactions that respond sensitively to changes in both internal and external environments. Particularly, the ovarian microenvironment acts as a critical biological foundation that orchestrates a series of processes ranging from follicular growth, oocyte maturation, and ovulation to fertilization and subsequent early embryonic development. Oocytes grow within the ovary through intimate interactions with various cells such as the granulosa and theca cells. The nutritional, metabolic, and signaling support provided by these cells is a crucial factor in determining oocyte quality and subsequent embryonic developmental competence (Orisaka et al., 2009; Turathum et al., 2021). Consequently, the impairment of homeostasis in the ovarian microenvironment extends beyond the follicular stage and reduces fertilization capacity, impairs early cleavage, and lowers blastocyst formation rates, ultimately leading to an overall decline in reproductive potential.
Recently, inflammatory responses within the ovarian microenvironment have consistently gained attention in the field of female reproductive study because of their close association with reproductive dysfunction (Boots & Jungheim, 2015; Snider & Wood, 2019; Ameho & Klutstein, 2025; Long et al., 2025). Fundamentally, inflammation is a physiological defense mechanism against tissue damage or infection; however, excessive or persistent inflammation disrupts tissue homeostasis and impairs cellular functions. In the reproductive system, the overexpression of pro-inflammatory cytokines, increased oxidative stress, and metabolic imbalances in the local microenvironment contribute to follicular developmental disorders and diminished oocyte quality. Specifically, clinical and experimental studies related to assisted reproductive technology have suggested that inflammatory conditions negatively impact fertilization rates, embryonic development rates, and implantation potential (Boots & Jungheim, 2015; Ameho & Klutstein, 2025). Therefore, elucidating the molecular and physiological changes associated with the inflammatory environment in the ovary and early embryos is critical for understanding the pathophysiology of reproductive disorders.
Follicular development is not merely a result of cell proliferation. It is a complex regulatory process involving continuous information exchange between the oocyte and surrounding somatic cells. Cumulus and granulosa cells support oocyte growth and maturation, whereas the oocyte regulates the functions of the surrounding cells to create an interdependent microenvironment (Orisaka et al., 2009; Appeltant et al., 2017). Therefore, in addition to inducing individual cellular changes, inflammatory stimuli within the ovarian microenvironment cause systemic dysfunction of the entire follicle by disrupting intercellular communication systems. This implies that the developmental competence of early embryos is not determined solely at the time of fertilization but is rather pre-established during the preceding stages of follicular formation and oocyte maturation. In this context, attempts to link and interpret inflammatory changes occurring at the ovarian level with impaired embryonic development are biologically well-founded.
In recent years, the relationship between inflammation and reproductive function has been increasingly substantiated through diverse experimental and clinical studies. Inflammation is partially involved in physiological processes such as ovulation, fertilization, and implantation. However, excessive or chronic activation of these responses impairs the homeostasis of reproductive organs (Boots & Jungheim, 2015; Snider & Wood, 2019; Ameho & Klutstein, 2025; Long et al., 2025). Overactive inflammatory responses within the ovarian tissue induce granulosa cell dysfunction, follicular atresia, increased apoptosis, and abnormal steroidogenesis, which potentially compromise the developmental competence of oocytes. Furthermore, key systemic inflammatory mediators and signaling receptors—such as tumor necrosis factor receptors (TNF-R) and macrophage colony-stimulating factor (M-CSF)—which are modulated during chronic inflammation, function as crucial factors that correlate with altered ovarian tissue homeostasis, cell viability, and the subsequent developmental competence of early embryos.
Lipopolysaccharide (LPS) is a component of the outer membrane of Gram-negative bacteria. It is an endotoxin that potently stimulates the innate immune system (Alexander & Rietschel, 2001). LPS activates inflammatory signaling pathways primarily through Toll-like receptor 4 (TLR4), which leads to the secretion of various cytokines and increased oxidative stress. Hence, LPS has been widely used to establish models of systemic or tissue-specific inflammation (Alexander & Rietschel, 2001; Andreasen et al., 2008). Furthermore, LPS-induced inflammation is frequently used in the study of the reproductive system for establishing experimental models for ovarian dysfunction, impaired oocyte maturation, and embryonic developmental abnormalities (Bromfield & Sheldon, 2011; Snider & Wood, 2019; Long et al., 2025). These models enable the evaluation of the manner in which inflammatory stimuli affect germ cells and their surrounding somatic cells. Notably, the ovary is an organ that sensitively reflects the impact of systemic inflammation owing to its robust blood supply and hormonal responsiveness. However, most studies have focused on the association between inflammatory responses and impaired follicular development, increased oxidative stress, and reduced fertilization rates (Bromfield & Sheldon, 2011; Boots & Jungheim, 2015; Snider & Wood, 2019; Ameho & Klutstein, 2025; Long et al., 2025). Consequently, relatively limited information is available on the mechanism by which inflammation induces changes in membrane lipid composition or glycosphingolipid metabolic pathways in the ovaries and embryos. Hence, although the fact that inflammation impairs reproductive function is relatively well established, the mechanism underlying the inflammation-mediated alteration of the lipid molecular networks remains insufficiently elucidated. This knowledge gap needs to be addressed to gain an in-depth understanding of inflammatory reproductive disorders.
Gangliosides are sialic acid-containing glycosphingolipids and major components of the cell membrane. They are involved in diverse biological functions such as intercellular recognition, receptor signaling, cell proliferation, differentiation, and maintenance of viability (Groux-Degroote et al., 2017; Sipione et al., 2020). Specifically, the lipid raft structure of the cell membrane significantly influences the spatial arrangement and functional interactions of receptor proteins and signaling molecules, and gangliosides are key molecules that regulate the characteristics of these membrane microdomains (Ryu et al., 2025). Therefore, alterations in ganglioside composition do not only indicate fluctuations in membrane components; instead, it underlies significant biological events that alter the manner in which cells respond to external stimuli.
Among the various gangliosides, GM3 exhibits the most fundamental structure and functions as an essential precursor for the biosynthesis of more complex gangliosides (Groux-Degroote et al., 2017; Sipione et al., 2020). GM3 biosynthesis is mediated by ST3GAL5, which is also known as GM3 synthase. Thus, alterations in the expression and activity of this enzyme are likely linked to changes in the accumulation of GM3 within the membrane environment. Gangliosides possess functional significance in various physiological and pathological phenomena including stem cell differentiation, nervous system development, and tumor cell signaling (Yoshida et al., 2001; Kwak et al., 2006; Groux-Degroote et al., 2017; Sipione et al., 2020). However, the roles of GM3 and ST3GAL5 in the female reproductive system—specifically in the ovarian microenvironment and during early embryonic development—remain largely uncharacterized in the field of reproductive biology.
Follicular and early embryonic development rely on diverse signaling systems based on cell membrane receptors. Growth factor receptors, cell adhesion molecules, and metabolism-related membrane proteins function in close association with the surrounding membrane lipid environment. In this context, changes in glycosphingolipid composition affect overall cellular function. Therefore, the disruption of ST3GAL5 expression and GM3-related metabolism in an inflammatory environment may be associated with alterations in membrane-based signaling networks that regulate oocyte maturation and embryonic development. Exploring the interactions between inflammatory responses and cell membrane ganglioside pathways may provide crucial clues for understanding the molecular background of impaired early embryonic development.
Melatonin (MT) is a well-known circadian rhythm-regulating hormone secreted by the pineal gland. MT has recently gained significant attention as a multifaceted molecule with simultaneous antioxidant, anti-inflammatory, and cytoprotective functions (Altun & Ugur-Altun, 2007; Radogna et al., 2010; Xia et al., 2012; Mauriz et al., 2013; Zhang & Zhang, 2014; Tan et al., 2015; Reiter et al., 2016; Kopustinskiene & Bernatoniene, 2021). MT mitigates tissue damage through various pathways involving reactive oxygen species (ROS) scavenging, inflammatory transcription factor regulation, mitochondrial function preservation, and apoptosis inhibition (Radogna et al., 2010; Mauriz et al., 2013; Zhang & Zhang, 2014; Reiter et al., 2016; Kopustinskiene & Bernatoniene, 2021). Furthermore, studies on ovarian function maintenance, oocyte quality improvement, and embryo protection have suggested that MT exerts beneficial effects; hence, it is a candidate of high interest for mitigating inflammatory reproductive microenvironments (Xia et al., 2012; Mauriz et al., 2013; Zhang & Zhang, 2014; Reiter et al., 2016; Kopustinskiene & Bernatoniene, 2021).
Nevertheless, whether MT protects reproductive function in inflammatory environments solely through its antioxidant actions or by influencing specific molecular pathways linked to cell membranes and lipid metabolism remains unclear. Particularly, extremely limited information is available on the interaction between ganglioside metabolism and MT and its potential relevance to ovarian function and early embryonic development. Therefore, elucidating the molecular axes through which MT exerts its protective effects against inflammation-induced reproductive damage is necessary for a precise understanding of its mechanisms.
The aim of the present study was to analyze the phenotypic changes in the developmental competence of the ovary and early embryos under LPS-induced chronic inflammation and observe corresponding alterations in GM3-related molecular alterations using an Institute of Cancer Research (ICR) mouse model. Specifically, the effect of inflammatory conditions on oocyte recovery and blastocyst formation rates was evaluated through superovulation and in vitro fertilization (IVF), while tracking parallel changes in ST3GAL5 expression in ovarian tissues and embryos. Furthermore, we examined whether these inflammation-induced reproductive impairments and biochemical alterations were alleviated by MT treatment to explore the potential correlation between inflammation, ganglioside metabolism, and early embryonic development. This study is significant because it expands the existing understanding of the inflammatory reproductive microenvironment and examines whether disruptions in GM3-related metabolism can serve as a molecular indicator associated with impaired early embryonic development.
MATERIALS AND METHODS
Ovarian tissues were collected, washed with phosphate-buffered saline (PBS), and homogenized under chilled conditions. Gangliosides were extracted according to previously described methods (Ladisch & Gillard, 1985) and isolated on a DEAE-Sephadex A-25 column (Sigma-Aldrich, St. Louis, MO, USA). The column was pre-washed with a mixture of chloroform/methanol/H2O (15:30:4, v/v), and acidic lipids were eluted using a solution of chloroform/methanol/0.8 M sodium acetate (15:30:4, v/v).
The eluted samples were dried under nitrogen gas at 30°C for 6 h. Then, they were dissolved in a mixture of 12 N NH4OH and chloroform/methanol (1:1, v/v) and allowed to react at room temperature for 16 h, followed by drying step under nitrogen gas. Next, the samples were reconstituted in 1 mL of distilled water and desalted using a Sep-Pak C18 cartridge (Millipore Sigma, Burlington, MA, USA). The gangliosides were eluted using methanol (2 mL) and chloroform/methanol (2:1, v/v, 4 mL), dried under nitrogen gas at 30°C for 4 h, and stored at −80°C until needed for further analysis.
The gangliosides were analyzed using HPTLC plates (5651; Merck, Darmstadt, Germany). Purified ganglioside samples (10 μL) were spotted onto plates using microhematocrit capillary tubes. The mobile phase comprised a chloroform/methanol/0.25% CaCl2 mixture (50:40:10, v/v/v). The gangliosides were identified by comparing with standard reference materials (Matreya LLC, State College, PA, USA) and visualized via resorcinol staining. For semi-quantitative densitometric analysis, the stained HPTLC plates were digitally scanned, and the band intensities were quantified using ImageJ software (National Institutes of Health, Bethesda, MD, USA). The background signal for each lane was subtracted to ensure measurement accuracy. The relative band intensity of ovarian GM3 was calculated by normalizing the value of each experimental group to that of the control group, which was arbitrarily set to 1.0.
In total, 60 ICR mice (6-week-old; 40 females and 20 males) were purchased from Samtako Bio Korea (Osan, Korea). The average body weight at the beginning of the experiment was 26.97±1.47 g. All mice were subjected to a one-week acclimation period to the laboratory environment prior to the study. The animals were maintained in a specific pathogen-free facility under a 12-h light/dark cycle. The temperature was maintained at 22±2°C and humidity at 50±10%. The mice were housed in groups of five per polycarbonate cage and provided Altromin Mouse Diet 1314 FORTIL (Altromin Spezialfutter GmbH, Lage, Germany) and sterilized water ad libitum. SAM-GB-L (Samtako Bio) was used as bedding.
All procedures were performed in compliance with the Guidelines for the Care and Use of Laboratory Animals and approved by the Institutional Animal Care and Use Committee (IACUC) Wonkwang University (Approval No.: WKU25-73; Approved on November 10, 2025).
To evaluate the downstream reproductive competence and parallel biochemical alterations established after sequential inflammatory exposure and pharmacological intervention, 7-week-old female ICR mice (n=40) were randomly assigned to four experimental groups (n=10 per group): Control group: Administered an equal volume of sterile PBS as a vehicle control following the same schedule. LPS group: Administered LPS (L4391, Sigma-Aldrich) via intraperitoneal (i.p.) injection at a dose of 10 mg/kg once every two days for 4 weeks to establish a history of chronic systemic inflammatory challenge. MT group: Administered melatonin (MT; M-5250, Sigma-Aldrich) via i.p. injection at a dose of 50 mg/kg once every two days for 4 weeks. LPS+MT group: Administered LPS (10 mg/kg, i.p.) once every two days during the first 2 weeks to induce an initial inflammatory state, followed by MT administration (50 mg/kg, i.p.) once every two days for the subsequent 2 weeks. This sequential regimen in the LPS+MT group was specifically structured to track the post-inflammatory recovery pattern during sequential LPS and melatonin exposure following a predefined period of prior inflammatory exposure, rather than assessing a preventive effect under continuous LPS challenge. However, because the administration of LPS was discontinued after the first 2 weeks, this experimental window evaluates the combined downstream phenotypes of pharmacological intervention alongside the potential natural clearance/withdrawal of the inflammatory stimulus. All dosages were strictly adjusted based on individual body weights measured prior to each injection. The male mice (n=20) were reared under identical environmental conditions and utilized solely for mature sperm collection for subsequent IVF assays. At the conclusion of the 4-week experimental period, all mice were humanely euthanized via cervical dislocation. Blood samples were immediately collected via cardiac puncture, and serum was isolated through centrifugation to evaluate the downstream alterations of systemic inflammatory profiles— specifically targeting key inflammatory markers and receptors including TNF-R1, TNF-R2, and M-CSF—via a serum cytokine antibody array kit. Concurrently, ovarian tissues and oocytes were harvested to evaluate the subsequent developmental competence and accompanying ganglioside metabolic profiles.
To induce superovulation in 11-week-old female mice, 10 IU of pregnant mare serum gonadotropin (Daesung Microbiological Labs, Uiwang, Korea) was administered intraperitoneally. After 48 h, 10 IU of human chorionic gonadotropin (hCG; Daesung Microbiological Labs) was injected via the same route. Oocytes were collected from the ampulla 16 h post-hCG injection for IVF. Spermatozoa obtained from male mice were suspended in human tubal fluid medium (HTF, MR-070; Sigma-Aldrich) and capacitated for 30 min at 37°C under 5% CO2. The capacitated sperm were co-incubated with the oocytes in HTF medium for 6 h. The resulting zygotes were cultured in potassium simplex optimized medium (MR-121; Sigma-Aldrich) at 37°C under 5% CO2. The embryos reached the blastocyst stage approximately 4.5 days after fertilization.
Ovarian tissues were harvested 6.5 h after hCG administration. The ovaries were fixed in 4% formalin (Sigma-Aldrich), dehydrated in a 30% sucrose solution, and embedded in OCT compound. Frozen sections (5-μm thick) were prepared using a cryostat and mounted onto coated slide glasses. The slides were air-dried for 1 min at room temperature (22±2°C), immersed in 70% ethanol at –20°C for 5 min, and fixed with 100% ethanol, followed by three 5-min washes in PBS. For permeabilization, the sections were treated with 0.1% Triton X-100 (Sigma-Aldrich) for 5 min and washed thrice with PBS. To prevent nonspecific binding, the slides were blocked with 1% bovine serum albumin for 30 min at room temperature (22±2°C). Then, the sections were incubated overnight at 4°C with a GM3 synthase antibody (B-12; sc-365329, 1:200; Santa Cruz Biotechnology, Dallas, TX, USA). After washing with PBS, the slides were incubated with a secondary antibody (Goat anti-Mouse IgG (H+L) Cross-Adsorbed Secondary Antibody, Alexa Fluor™ 488; A-11001, 1:500; Thermo Fisher Scientific, Waltham, MA, USA) in the dark for 1 h at room temperature (22±2°C). The nuclei were counterstained with DAPI (Thermo Fisher Scientific) for 5 min in the dark, followed by three 5-min PBS washes. Finally, the slides were mounted with anti-fade mounting medium and visualized under a fluorescence microscope (ZEISS, Oberkochen, Germany) at 100× magnification. For the semi-quantitative analysis of GM3 synthase expression, the mean fluorescence intensity was measured using ImageJ software (National Institutes of Health). Specific regions of interest were selected under identical exposure and threshold settings for all samples, and the non-specific background fluorescence was subtracted from each measurement. The relative fluorescence intensity was calculated by normalizing the quantified values to that of the control group.
Cytokine profiling was performed using a Mouse Cytokine Array C3 Kit (RayBiotech, Norcross, GA, USA) according to the manufacturer’s instructions. Briefly, serum was separated from the blood samples by centrifuging at 9,358×g for 5 min. The resulting supernatant was 2–5-fold diluted with blocking buffer. The membranes were incubated with blocking buffer at 20 rpm ([LK Labkorea] Digital Shaker) for 30 min for stabilization. Subsequently, the diluted samples were applied to the membranes and incubated at 4°C for at least 16 h. Then, the membranes were washed thrice with 1× Washing Buffer I and twice with 1× Washing Buffer II for 5 min each. A biotinylated antibody cocktail was mixed with the blocking buffer and applied to the membranes, followed by overnight incubation at 4°C. The same washing procedure was performed, after which horseradish peroxidase (HRP)-conjugated streptavidin diluted to a 1× concentration was added. The membranes were incubated overnight at 4°C. After a final wash, the detection solution was prepared by mixing the Detection Buffers A and B. This mixture was applied to the membranes, and the signals were captured using chemiluminescence detection. For the quantitative profiling of cytokine levels, the pixel density of each dot on the membranes was analyzed by densitometry using ImageJ software (National Institutes of Health). The background signal surrounding each spot was subtracted from the measured value. The relative expression levels of individual cytokines were calculated by normalizing the spot intensities to the positive control spots on the membrane or the experimental control group.
Band, spot, and fluorescence intensities were quantified by densitometric analysis using ImageJ software (v1.53; National Institutes of Health). All quantitative data from independent replicates were analyzed using GraphPad Prism software (v9.0; GraphPad Software, San Diego, CA, USA) and are presented as the mean±SD. Statistical significance among multiple groups was determined using one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test. Statistical significance is indicated by asterisks (* p<0.05) or ‘ns’ for non-significant differences. Data represent a descriptive semi-quantitative assessment from a single representative analysis, and inferential statistical testing was not applied.
RESULTS
The results of the HPTLC analysis of gangliosides extracted from ovarian tissues are presented in Fig. 1. In the control group, the GM3 band was observed with high clarity, reflecting the baseline GM3 abundance in normal ovarian tissue. In contrast, the relative intensity of the GM3 band was lower in the LPS-treated group, indicating that prior exposure to chronic inflammatory stimuli is associated with a reduction in GM3 levels within the ovaries. These changes suggest that inflammation-associated stress correlates not only with systemic responses but also with alterations in the membrane lipid environment of ovarian tissues.

In the LPS+MT group, the GM3 signal showed a partial restorative trend compared to the diminished level in the LPS-only group. This result provides biochemical evidence that subsequent melatonin intervention is accompanied by a moderate stabilization of impaired ganglioside profiles in the ovaries following inflammatory challenge. Consequently, the relative alterations shown in Fig. 1 serve as primary data that parallel the subsequent embryonic development outcomes and immunofluorescence staining patterns.
We evaluated the impact of inflammatory conditions on downstream reproductive outcomes by performing IVF after superovulation in each experimental group, comparing the total number of recovered oocytes and blastocyst formation rates (Fig. 2). In total, 121 oocytes were recovered from the control group, of which 69.4% developed normally to the blastocyst stage, indicating that baseline oocyte production and early embryonic developmental competence were maintained in our experimental system. In contrast, the number of recovered oocytes dropped to 74 in the LPS-treated group, and the blastocyst formation rate decreased to 33.7%, suggesting that the prior inflammatory challenge adversely affected both the follicular environment and subsequent early embryonic development stages.
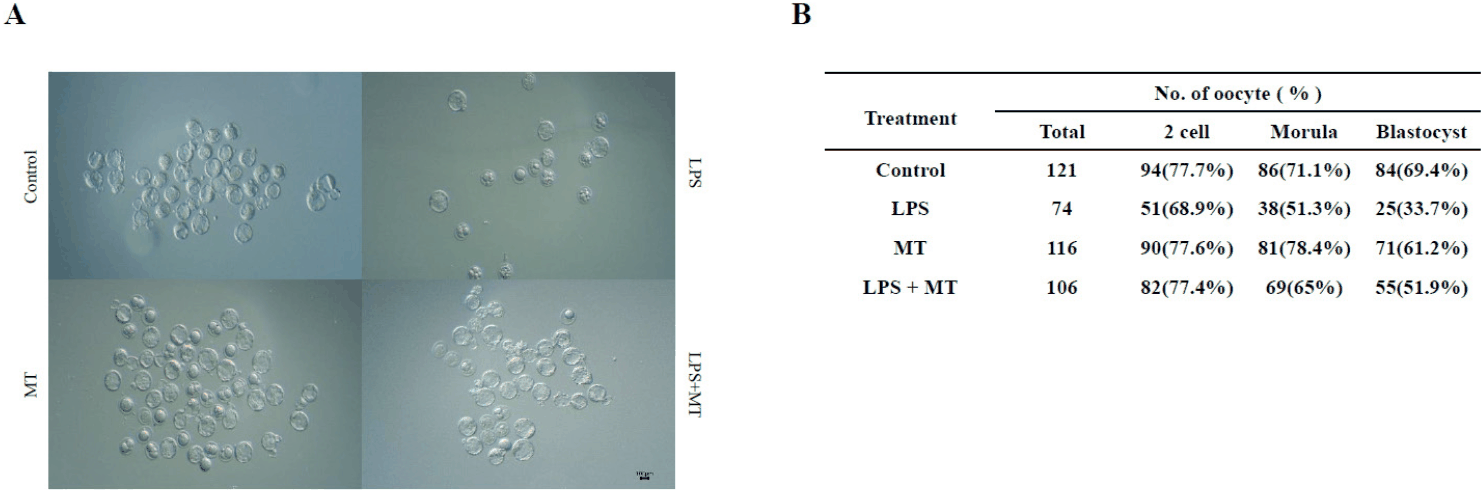
In the LPS+MT group, the total number of recovered oocytes was 106, and the blastocyst formation rate was observed at 51.9%. Although these values did not fully reach those of the control group, a distinct partial recovery trend was observed in both oocyte recovery and blastocyst formation compared with the LPS-only group. These results indicate that LPS-induced inflammation was associated with diminished reproductive outcomes and that melatonin intervention partially mitigates this decline.
Changes in systemic inflammatory responses were analyzed using a serum cytokine antibody array. The results are presented in Fig. 3. Compared with the control group, the LPS-treated group exhibited increased signals for several inflammatory cytokines. Semi-quantitative analysis of representative cytokines (TNF-R1, TNF-R2, and M-CSF) using ImageJ showed higher relative expression levels in the LPS-treated group than in the control group (Fig. 3C–E).
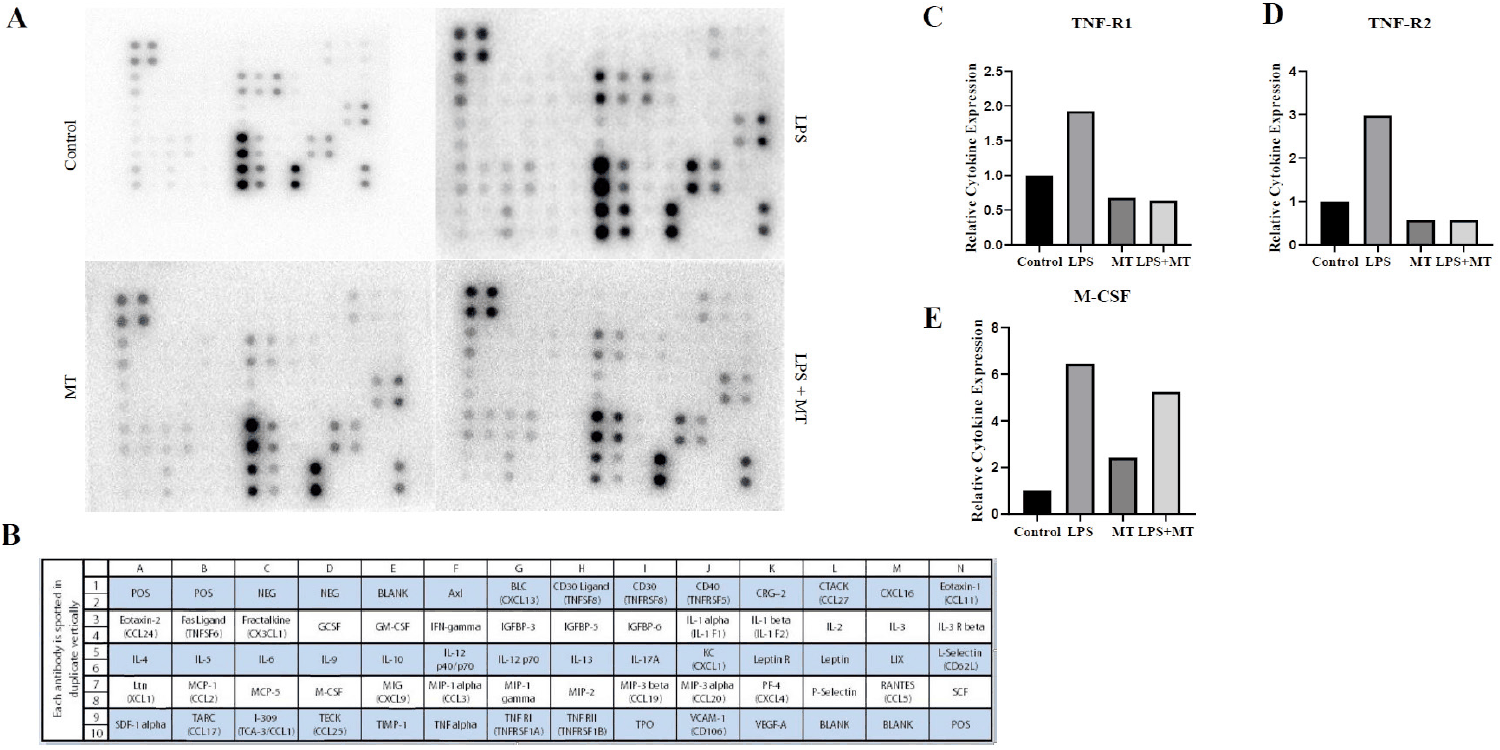
In contrast, the elevated cytokine signals observed in the LPS-treated group showed a partial reduction trend in the LPS+MT group, whereas the MT-only group exhibited expression patterns comparable to those of the control group. Relative expression levels were normalized to the control group (set to 1.0) to present a descriptive comparison of serum profiles. These results indicate differences in serum inflammatory cytokine expression among the experimental groups.
The immunofluorescence staining and corresponding semi-quantitative analyses for ST3GAL5 in blastocyst-stage embryos are shown in Fig. 4. In the control blastocysts, the embryos uniformly exhibited ST3GAL5 (GM3 synthase) signals with relative clarity, indicating that ST3GAL5 protein is distributed during normal embryonic stages (Fig. 4A; e, i). In contrast, the relative fluorescence signal was generally weaker in the LPS-treated group (Fig. 4A; f, j), suggesting that prior maternal inflammatory exposure correlates with a downregulation of ganglioside-related enzyme distribution at the early embryonic stage.
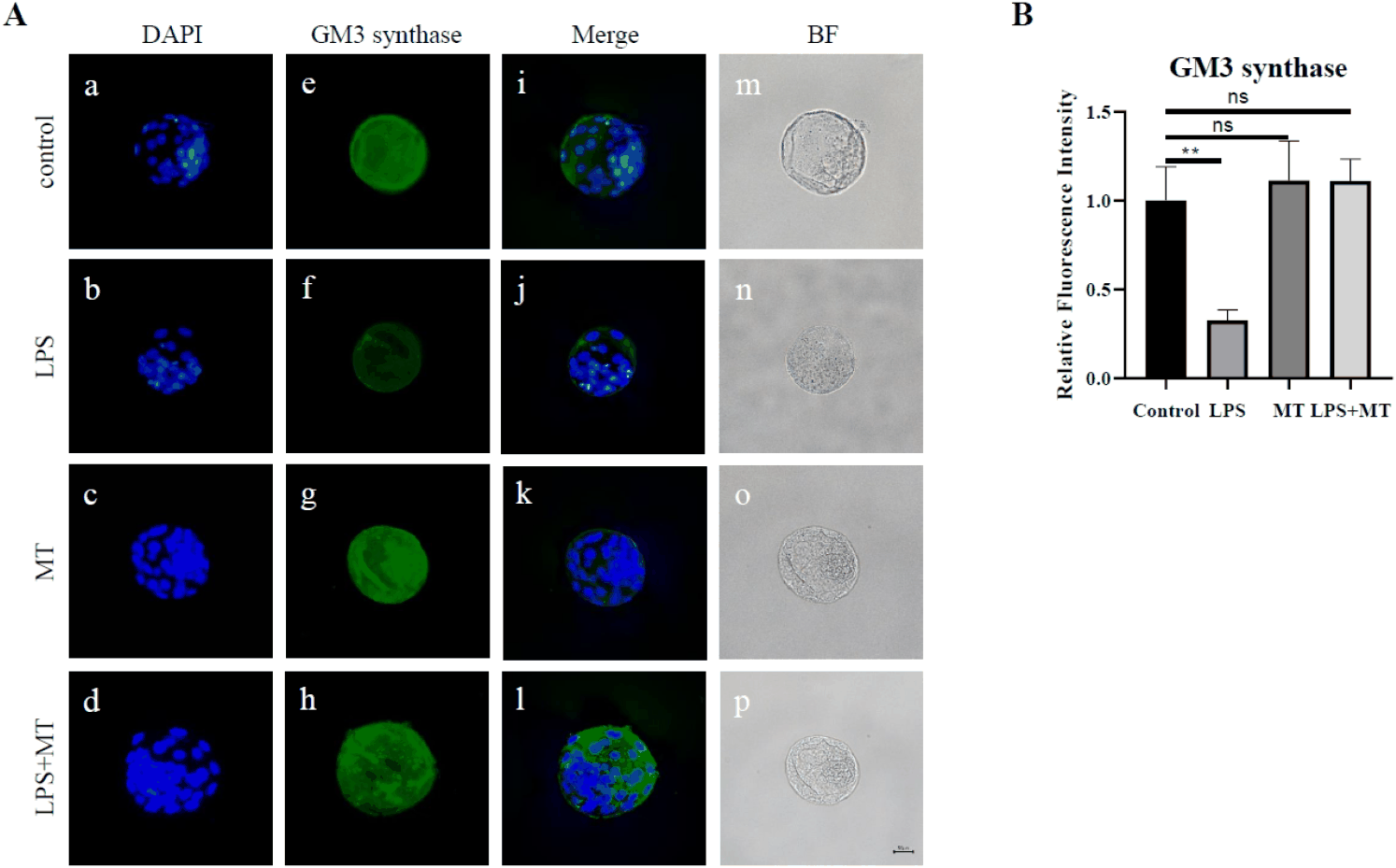
These biochemical changes are noteworthy when interpreted along with the reduction in blastocyst formation rates (Fig. 2), implying that maternal LPS exposure correlates not merely with a lower rate of embryonic development but also with parallel alterations in the molecular profile of the embryos. Importantly, semi-quantitative graphical analysis of the relative fluorescence intensity confirmed that subsequent melatonin administration in the LPS+MT group was accompanied by a partial recovery trend in the ST3GAL5 signal compared to the group exposed to LPS alone (Fig. 4B). These data indicate that partial improvement in developmental competence after melatonin treatment occurred in parallel with partial recovery of ST3GAL5 fluorescence; however, a causal relationship between these changes was not established.
Finally, Fig. 5 illustrates the immunofluorescence distribution of ST3GAL5 across various follicular developmental stages in the ovarian tissue, accompanied by detailed semi-quantitative comparisons across specific follicular cell types. In the control group, ST3GAL5 localization was identified across multiple stages, including primary, secondary, and Graafian follicles, with distinct relative signals observed particularly in the theca and granulosa cells. This suggests that GM3-related profiles persist throughout follicular development and may be associated with the maintenance of the baseline ovarian microenvironment.
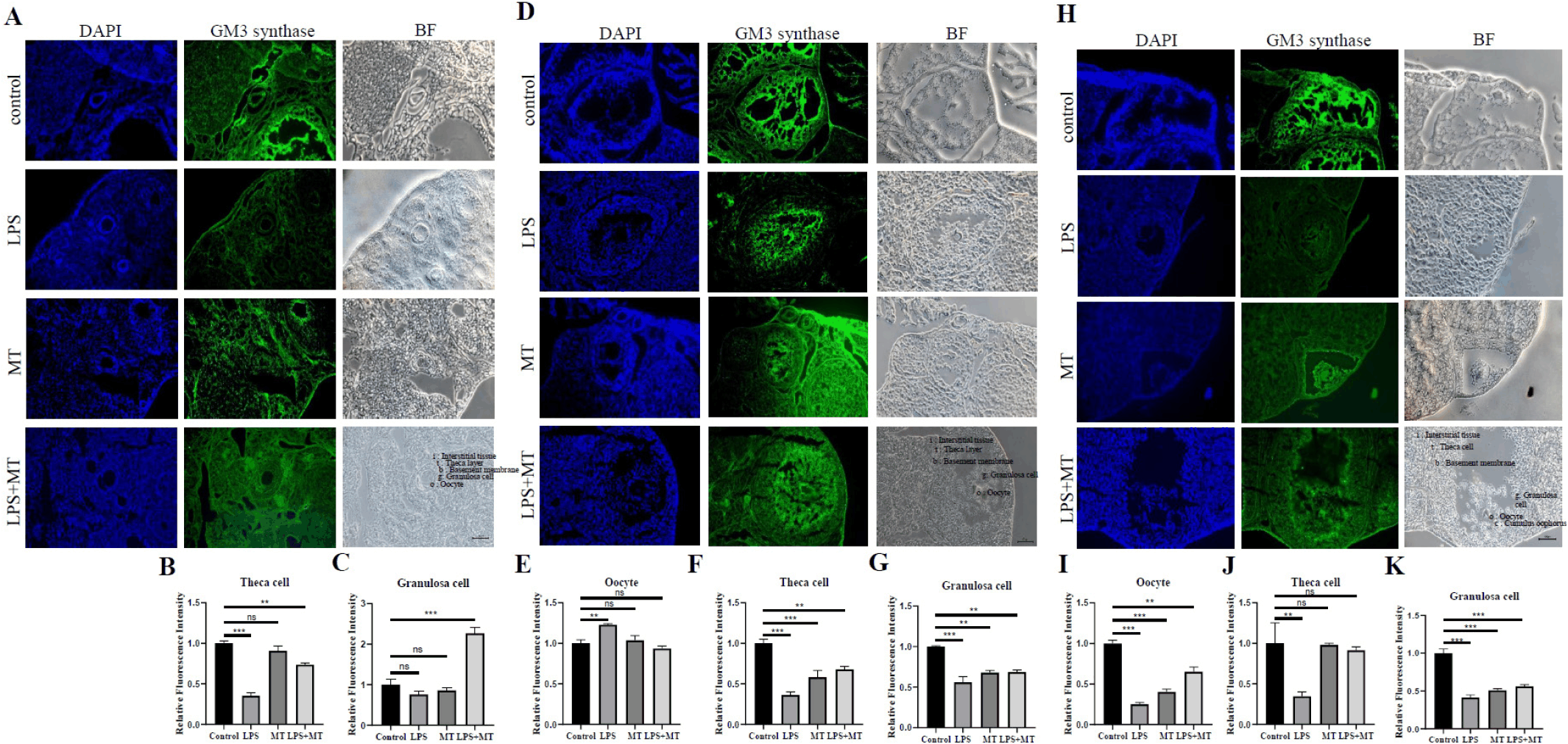
In contrast, weak ST3GAL5 signals were observed across the ovarian structures in the LPS-treated group, whereas this reduction was partially restored in the LPS+MT group. Specifically, semi-quantitative graphical analysis revealed that prior exposure to LPS was associated with a noticeable decrease in relative staining intensity within primary follicle layers (Fig. 5A–C), secondary follicle components (Fig. 5D–G), and mature Graafian follicles (Fig. 5H–K). Subsequent melatonin intervention in the LPS+MT group was associated with a moderate elevation and partial recovery trend in the relative staining intensity across these respective somatic and germ cell layers (Fig. 5B, C, E–G, I–K).
These immunohistochemical observations complement and support the HPTLC findings shown in Fig. 1. While Fig. 1 indicates a biochemical decline in total GM3 levels within the ovary, the spatial tracking in Fig. 5 confirms that these alterations occur consistently at the morphological level within specific follicular structures. Therefore, these findings suggest that variations in the ST3GAL5-associated membrane environment occur concurrently with inflammation-induced reproductive impairment and its subsequent pharmacological modulation.
DISCUSSION
The present study demonstrated that LPS-induced chronic inflammation is associated with alterations in the ovarian microenvironment and impaired early embryonic development in ICR mice. Notably, these changes were accompanied by a reduction in GM3 levels (Fig. 1) and a downregulation of ST3GAL5 expression (Figs. 4 and 5) in both ovarian tissues and blastocysts.
Compared with the control group, the LPS-treated group exhibited marked reductions in both the number of retrieved oocytes and the blastocyst formation rate (Fig. 2). These findings align with previous reports indicating that an inflammatory environment is associated with a decline in female reproductive function and embryonic developmental competence (Boots & Jungheim, 2015; Snider & Wood, 2019; Ameho & Klutstein, 2025; Long et al., 2025). Given the intimate bidirectional communication between granulosa cells and oocytes during follicular development (Orisaka et al., 2009), our results suggest that inflammation-induced alterations in the ovarian microenvironment potentially contribute to compromised oocyte quality and subsequent reduction in developmental competence.
LPS is widely utilized as a standard experimental agent to induce inflammation by activating innate immune signaling pathways and stimulating the production of various inflammatory mediators (Alexander & Rietschel, 2001; Andreasen et al., 2008; Bromfield & Sheldon, 2011). In this study, cytokine antibody array analysis revealed distinct shifts in serum inflammatory cytokine profiles in the LPS-treated group compared to the control group (Fig. 3). Semi-quantitative analysis further confirmed these alterations in key cytokines, including TNF-R1, TNF-R2, and M-CSF. Because inflammatory cytokines are known to influence ovarian homeostasis, oxidative stress, and intercellular signaling within reproductive tissues, the observed declines in retrieved oocyte numbers and blastocyst formation rates may represent a complex, multifaceted outcome linked to the disrupted ovarian microenvironment rather than the consequence of a single downstream signaling pathway.
A distinctive feature of this study is the evaluation of inflammation-induced reproductive decline in conjunction with molecular changes related to gangliosides. Gangliosides are glycosphingolipids that play pivotal roles in maintaining cell membrane structural stability and modulating receptor-mediated signaling (Groux-Degroote et al., 2017; Sipione et al., 2020). Among these, GM3 serves as a biosynthetic precursor for more complex gangliosides and is synthesized by ST3GAL5 (Ryu et al., 2025). We observed a decrease in ST3GAL5 expression in both ovarian tissues (Fig. 5) and blastocysts (Fig. 4). These findings suggest a potential link between an inflammatory environment and altered expression of GM3-related molecules during early embryonic development. However, since this study relied primarily on HPTLC and semi-quantitative immunofluorescence analysis, these results should be interpreted as demonstrating an association between GM3 metabolism and reproductive decline rather than establishing a direct causal mechanism.
Follicular development and early embryonic growth are highly sophisticated processes requiring precise regulation of cell proliferation, differentiation, intercellular adhesion, and receptor-mediated signaling. Alterations in the glycosphingolipid composition of cell membranes can modify the membrane-based molecular environment that governs these biological events. Previous studies have underscored the critical roles of gangliosides in stem cell differentiation, nervous system development, and cellular signaling regulation (Yoshida et al., 2001; Kwak et al., 2006; Groux-Degroote et al., 2017; Sipione et al., 2020; Ryu et al., 2025). Although the observed alterations under inflammatory conditions underscore a correlation between inflammation and altered ST3GAL5 protein signals, the specific downstream pathways were not investigated in this study. Thus, our findings should be viewed as highlighting an association between an inflammatory milieu and ST3GAL5 expression rather than proving a direct functional role of GM3 synthase in early embryonic developmental arrest.
Interestingly, the LPS+MT group showed improvements in both the number of retrieved oocytes and the blastocyst formation rate compared to the LPS-alone group (Fig. 2). Furthermore, a partial recovery trend was observed in serum inflammatory cytokine expression (Fig. 3) and ST3GAL5 expression in both ovarian tissues (Fig. 5) and blastocysts (Fig. 4). These observations are generally consistent with prior studies reporting the antioxidant and anti-inflammatory potential associated with melatonin (MT) (Altun & Ugur-Altun, 2007; Radogna et al., 2010; Xia et al., 2012; Mauriz et al., 2013; Zhang & Zhang, 2014; Tan et al., 2015; Reiter et al., 2016; Kopustinskiene & Bernatoniene, 2021). Nevertheless, these results indicate an association between MT treatment and these restorative changes, without definitively proving direct causality or a specific mechanism of action.
Consequently, a cautious approach is warranted when interpreting the effects of MT. Although the LPS+MT group demonstrated improvements in reproductive parameters compared to the LPS group, not all evaluated indices fully recovered to control levels. Moreover, our experimental design utilized a sequential treatment regimen: LPS was administered during the first two weeks and then discontinued, followed by MT administration for the subsequent two weeks. Therefore, the observed restorative changes may reflect a combination of the effects of MT treatment and the natural endogenous recovery process that occurred after the inflammatory stimulus was removed. Because this study did not include an experimental group subjected to continuous LPS administration alongside MT, we could not clearly differentiate the independent pharmacological effects of MT from the body’s intrinsic recovery mechanisms. Accordingly, these findings should be understood as indicating a partial recovery pattern following prior exposure to inflammation rather than establishing a definitive mitigating or preventive action of MT.
The molecular link between MT treatment and changes in ST3GAL5 expression warrants further investigation. MT is known to participate in diverse biological processes, including scavenging ROS, modulating inflammation-related transcription factors, maintaining mitochondrial function, and inhibiting apoptosis (Radogna et al., 2010; Mauriz et al., 2013; Zhang & Zhang, 2014; Reiter et al., 2016; Kopustinskiene & Bernatoniene, 2021). However, because these pathways were not directly analyzed in this study, it remains unclear whether the observed changes in ST3GAL5 expression resulted from a direct action of MT or were secondary effects of an overall improvement in the inflammatory microenvironment. Future studies should comprehensively analyze oxidative stress markers, mitochondrial function, inflammatory signaling pathways, and the transcriptional regulation of ST3GAL5 to elucidate these interrelationships more clearly.
The academic significance of this study lies in its evaluation of inflammation-induced reproductive decline from the perspective of ganglioside-related molecular alterations alongside reproductive outcomes. While existing literature has predominantly focused on oxidative stress, inflammatory cytokines, and compromised oocyte quality (Boots & Jungheim, 2015; Snider & Wood, 2019; Ameho & Klutstein, 2025; Long et al., 2025), this study analyzed ST3GAL5 expression in both ovarian tissues and blastocysts, highlighting the possibility that inflammation-linked GM3 molecular changes occur across early reproductive stages. Although these findings do not confirm a direct functional role for GM3 metabolism, they provide foundational data for future research aiming to clarify the relationship between ganglioside metabolism and inflammatory reproductive decline.
Several limitations of this study should be acknowledged. First, because this research was conducted using a mouse model, future validation in human or more clinically relevant experimental models is required. Second, since GM3-related alterations were evaluated mainly through HPTLC and semi-quantitative immunofluorescence, precise quantification of individual gangliosides was limited. Implementing LC-MS/MS-based ganglioside profiling in future studies would allow for a more rigorous molecular-level analysis. Third, the functional role of ST3GAL5 was not directly validated using gain- or loss-of-function approaches; hence, a causal relationship between the observed alterations and reproductive decline cannot be firmly established. Finally, the absence of an experimental group maintained under continuous LPS stimulation during MT administration precluded a clear differentiation between the pharmacological efficacy of MT and the relative contribution of natural recovery following the cessation of inflammation. Therefore, future investigations incorporating continuous inflammation models, ST3GAL5 functional modulation experiments, single-cell analysis, and human-based reproductive models are necessary to precisely elucidate the biological significance and clinical applicability of the molecular changes observed in this study.