
INTRODUCTION
Breast cancer is one of the most common malignancies and a leading cause of cancer-relatedmortality worldwide. Despite advances in early detection and treatment, tumor recurrence and metastasis remain major clinical challenges (Bray et al., 2018). Tumor metastasis is a complex process that involves enhanced migratory and invasive capabilities of cancer cells, which are closely associated with poor clinical outcomes (Bray et al., 2018).
Cancer stem cells (CSCs) have been proposed to play a critical role in tumor initiation, progression, and therapeutic resistance (Al-Hajj et al., 2003; Dean et al., 2005). CSCs possess self-renewal capacity and are characterized by the expression of specific markers, including CD44 and aldehyde dehydrogenase family members such as ALDH1A1 and ALDH1A3 (Al-Hajj et al., 2003; Ginestier et al., 2007). Accumulating evidence suggests that CSCs are also closely associated with metastatic potential (Dean et al., 2005; Zhang et al., 2023).
Epithelial-mesenchymal transition (EMT) is a biological process that enables epithelial cells to acquire mesenchymal properties, leading to increased migration and invasion (Thiery et al., 2009; Nieto et al., 2016). During EMT, epithelial markers such as E-cadherin are downregulated, whereas mesenchymal markers, including N-cadherin and vimentin, are upregulated, along with EMT-associated transcription factors such as Snail, Slug, Twist, and Zeb1 (Thiery et al., 2009). EMT has been shown to be closely linked to the acquisition of CSC-like characteristics and metastatic behavior (Mani et al., 2008).
Baculoviral inhibitor of apoptosis repeat containing 5 (BIRC5), also known as survivin, is a member of the inhibitor of apoptosis protein family and plays a key role in regulating cell survival and proliferation (Li et al., 1998; Altieri, 2008). BIRC5 is highly expressed in various cancers, including breast cancer, and has been associated with poor prognosis (Altieri, 2008; Jaiswal et al., 2015). Although previous studies have primarily focused on its role in apoptosis and tumor growth, its involvement in CSC regulation, EMT, and metastatic potential in breast cancer remains unclear (Altieri, 2008; Jaiswal et al., 2015).
In this study, we investigated the role of BIRC5 in regulating CSC-like properties, EMT, and migration/invasion abilities in human breast cancer cells. Using sphere-forming conditions and siRNA-mediated knockdown approaches, we examined the effects of BIRC5 on CSC marker expression, self-renewal capacity, EMT-related proteins, and metastatic behaviors. Our findings provide evidence that BIRC5 is involved in multiple processes associated with breast cancer progression.
MATERIALS AND METHODS
The human breast cancer cell line MDA-MB-231 (HTB-26) was obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). Cells were cultured in high-glucose Dulbecco’s modified Eagle’s medium (DMEM; LM001-05, Welgene, Gyeongsan, Korea) supplemented with 10% fetal bovine serum (FBS; 16000-044, Gibco, Thermo Fisher Scientific, Waltham, MA, USA) and 1% penicillin-streptomycin (SV30010, HyClone, Cytiva, Marlborough, MA, USA). Cells were maintained at 37°C in a humidified incubator with 5% CO2.
Primary antibodies used for western blot analysis were obtained from the following sources: Santa Cruz Biotechnology (Dallas, TX, USA): CD44 (sc-9960), ALDH1A1 (sc-374149), Sox2 (sc-365823), vimentin (sc-6260), Zeb1 (sc-25388), Slug (sc-166476), Snail (sc-271977), and Twist (sc-81417); Abcam (Cambridge, Abcam UK): ALDH1A3 (ab129815), Oct4 (ab181557), and BIRC5 (ab76424); Cell Signaling Technology (Danvers, MA, USA): Nanog (#4893); BD Transduction Laboratories (Franklin Lakes, NJ, USA): E-cadherin (610182) and N-cadherin (610920).
Cells were transfected with BIRC5-specific siRNA (sc-29499, Santa Cruz Biotechnology) using Lipofectamine RNAiMAX (13-778-150, Invitrogen, Thermo Fisher Scientific) according to the manufacturer’s instructions. After transfection, cells were incubated for 48 h at 37°C in a humidified atmosphere containing 5% CO2.
Kaplan-Meier survival curves were generated using the Kaplan-Meier plotter database (https://kmplot.com/analysis), based on mRNA expression data from breast cancer patients. BIRC5 expression was analyzed using all available patient samples with default parameters.
The mRNA expression level of BIRC5 in breast cancer was analyzed using the GEPIA2 database (http://gepia2.cancer-pku.cn), based on the Cancer Genome Atlas (TCGA) and GTEx datasets.
Total RNA was extracted using TRIzol reagent (TR118, Molecular Research Center, Cincinnati, OH, USA) according to the manufacturer’s instructions. RNA was precipitated with isopropanol (1.09634.1011, Merck KGaA, Darmstadt, Germany), washed with 75% ethanol (1.00983.1011, Merck KGaA), and dissolved in diethylpyrocarbonate (DEPC)-treated water. cDNA was synthesized using RT Premix (Random Primer; 25082, LiliF Diagnostics, Seoul, Korea). PCR amplification was performed using PCR PreMix (iMAX II; 25265, LiliF Diagnostics) with a Mastercycler Nexus Gradient system (Eppendorf, Hamburg, Germany). PCR products were separated on 2% agarose gels (50153, FMC BioProducts, Philadelphia, PA, USA) prepared in TAE buffer (IBSD-BT002, iNtRON Biotechnology, Seongnam, Korea) and visualized using a UV imaging system (MaestroGen, Hsinchu, Taiwan).
Cells were lysed in RIPA buffer (R4100-050, GenDEPOT, Katy, TX, USA) supplemented with a protease inhibitor cocktail (11836153001, Roche, Basel, Switzerland). Protein concentrations were determined using a Bradford assay (#5000006, Bio-Rad, Hercules, CA, USA). Equal amounts of protein were separated by SDS-PAGE and transferred onto nitrocellulose membranes. Membranes were incubated with primary antibodies overnight at 4°C, followed by HRP-conjugated secondary antibodies (anti-Rabbit: 7074s, anti-Mouse: 7076s, Cell Signaling Technology) for 1–2 h at room temperature. Protein bands were detected using enhanced chemiluminescence reagents (32106, Invitrogen, Thermo Fisher Scientific).
Cells were cultured in CSC-inducing conditioned medium (CSC-CM) to induce CSC-like properties. Cells were cultured in serum-free DMEM/F12 medium (11320-033, Gibco, Thermo Fisher Scientific) supplemented with basic fibroblast growth factor (bFGF, 20 ng/mL; 13256-029, Invitrogen, Thermo Fisher Scientific), epidermal growth factor (EGF, 20 ng/mL; E9644, Sigma-Aldrich, St. Louis, MO, USA), N-2 supplement (17502-048, Gibco), and B27 supplement (17504-044, Gibco). Cells were incubated for at least 3 days at 37°C in 5% CO2 to allow sphere formation.
For single cell assay, individual cells were seeded into 96-well plates (701003, Ushelf, Hopkinton, MA, USA) and cultured in CSC-CM. After 10–14 days, the number of spheres formed in each well was counted.
Cell migration assays were performed using uncoated Transwell chambers (3422, Corning, Corning, NY, USA). For invasion assays, chambers were coated with Matrigel (356234, Corning) according to the manufacturer’s instructions. Cells (1.5×104) suspended in serum free medium (Opti-MEM; 11058-021, Gibco, Thermo Fisher Scientific) were seeded into the upper chamber, while medium containing 10% FBS was added to the lower chamber. After 48 h incubation at 37°C, non-migrated cells were removed, and migrated or invaded cells were stained with crystal violet (HT90132-1L, Sigma-Aldrich) and counted under a light microscope.
Band intensities from Western blot and RT-PCR gel images were quantified by densitometric analysis using ImageJ software (National Institutes of Health, Bethesda, MD, USA). The intensity of each band was measured from the corresponding image, and relative expression levels were calculated by normalization to the control group. The control group was assigned a value of 1.0, and the values of the treatment groups were expressed as fold changes relative to the control.
All experiments were performed with at least three independent biological replicates. Statistical analyses were performed using Microsoft Excel (Microsoft Corporation, Redmond, WA, USA). Experimental data are presented as mean±SD from five or more independent experiments. The exact n value for each experiment is indicated in the corresponding figure legend. Comparisons between groups were performed using two-tailed paired Student’s t-tests or one-way analysis of variance (ANOVA). Not significant (ns): p>0.05; significant.
RESULTS
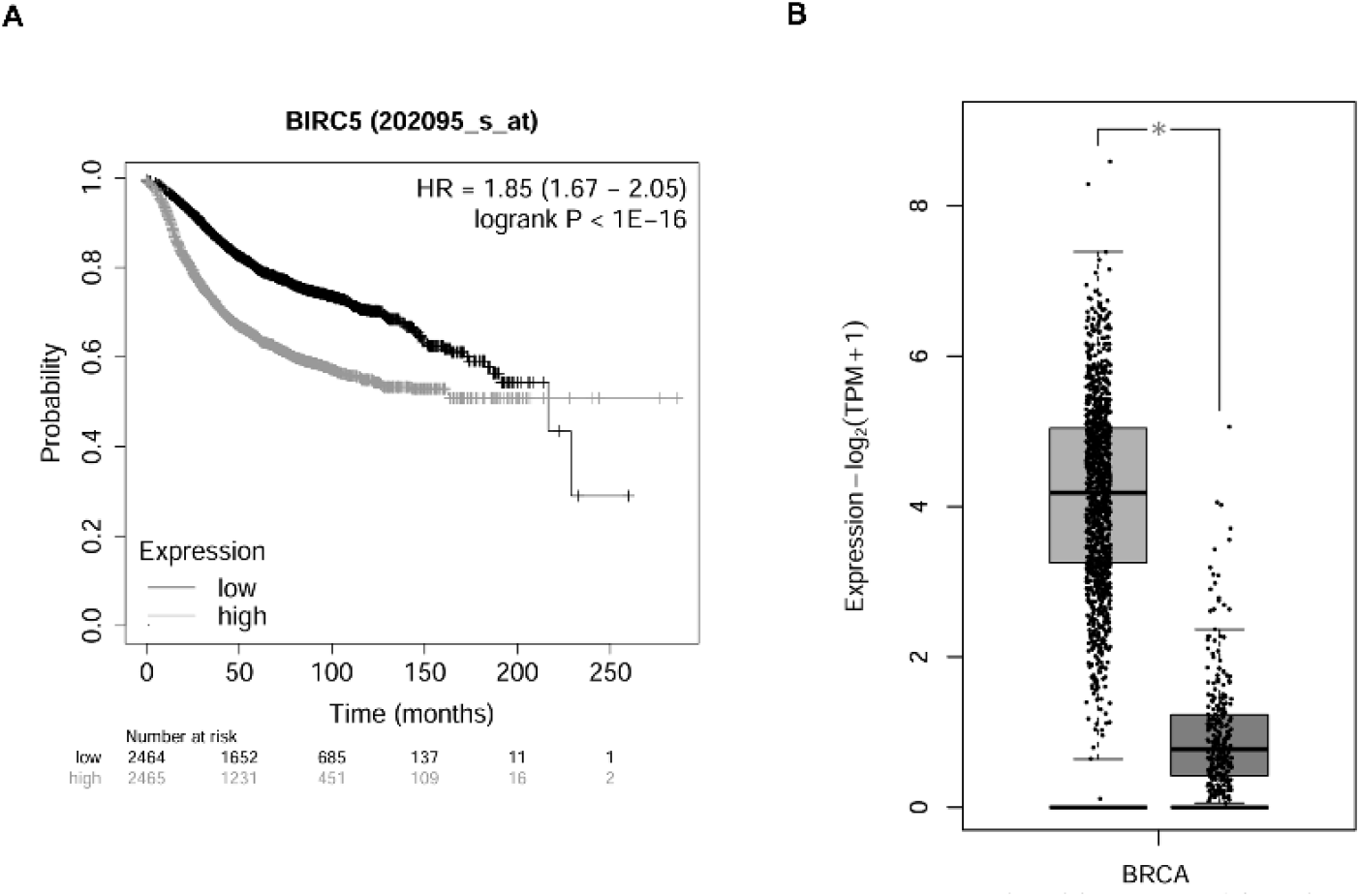
To investigate the clinical relevance of BIRC5, its expression and prognostic significance were analyzed using publicly available breast cancer datasets. Kaplan-Meier survival analysis demonstrated that patients with high BIRC5 expression exhibited significantly reduced overall survival compared with those with low expression (Fig. 1A). In addition, BIRC5 expression was significantly higher in breast cancer tissues than in normal tissues (Fig. 1B). These findings suggest that elevated BIRC5 expression is associated with poor clinical outcomes in breast cancer.
To determine whether BIRC5 is associated with CSC-like properties, breast cancer cells were cultured under sphere forming conditions. Western blot analysis revealed that sphere culture increased the protein expression levels of CSC markers, including CD44, ALDH1A1, and ALDH1A3. In addition, BIRC5 protein expression was also elevated under these conditions (Fig. 2A).
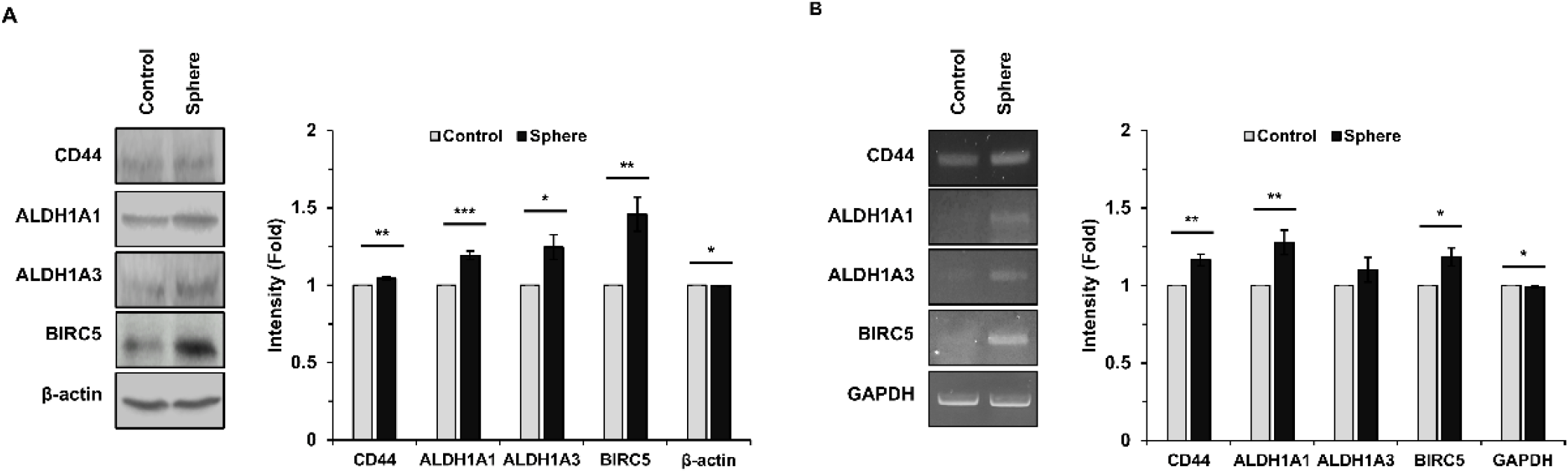
To further examine whether these changes occur at the transcriptional level, RT-PCR analysis was performed. Consistent with the protein expression data, mRNA levels of CD44, ALDH1A1, ALDH1A3, and BIRC5 were increased in cells cultured under sphere forming conditions compared with adherent control cells (Fig. 2B).
These results indicate that sphere forming conditions enhance the expression of CSC associated markers and BIRC5 at both protein and mRNA levels, suggesting a potential role of BIRC5 in CSC-like properties.
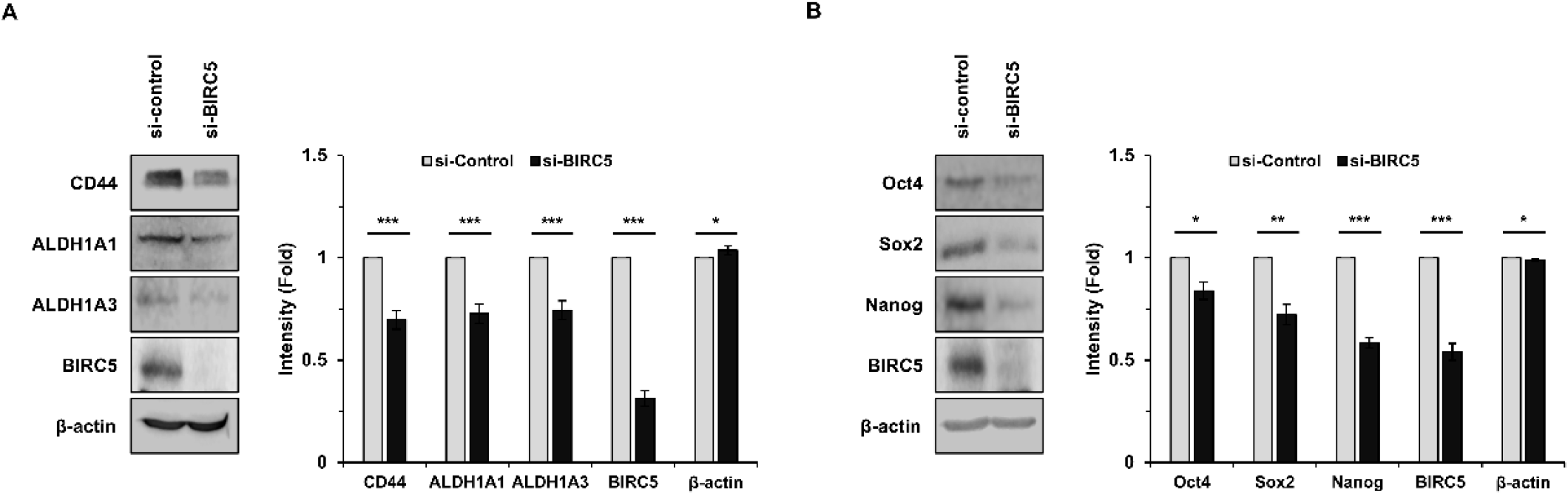
To further examine the role of BIRC5 in CSC regulation, BIRC5 expression was silenced using siRNA. Knockdown of BIRC5 markedly reduced the expression of CSC markers, including CD44, ALDH1A1, and ALDH1A3 (Fig. 3A). In addition, the expression of pluripotency-associated transcription factors, such as Sox2, Oct4, and Nanog, was also decreased following BIRC5 silencing (Fig. 3B). These findings suggest that BIRC5 is involved in the regulation of CSC-associated gene expression.
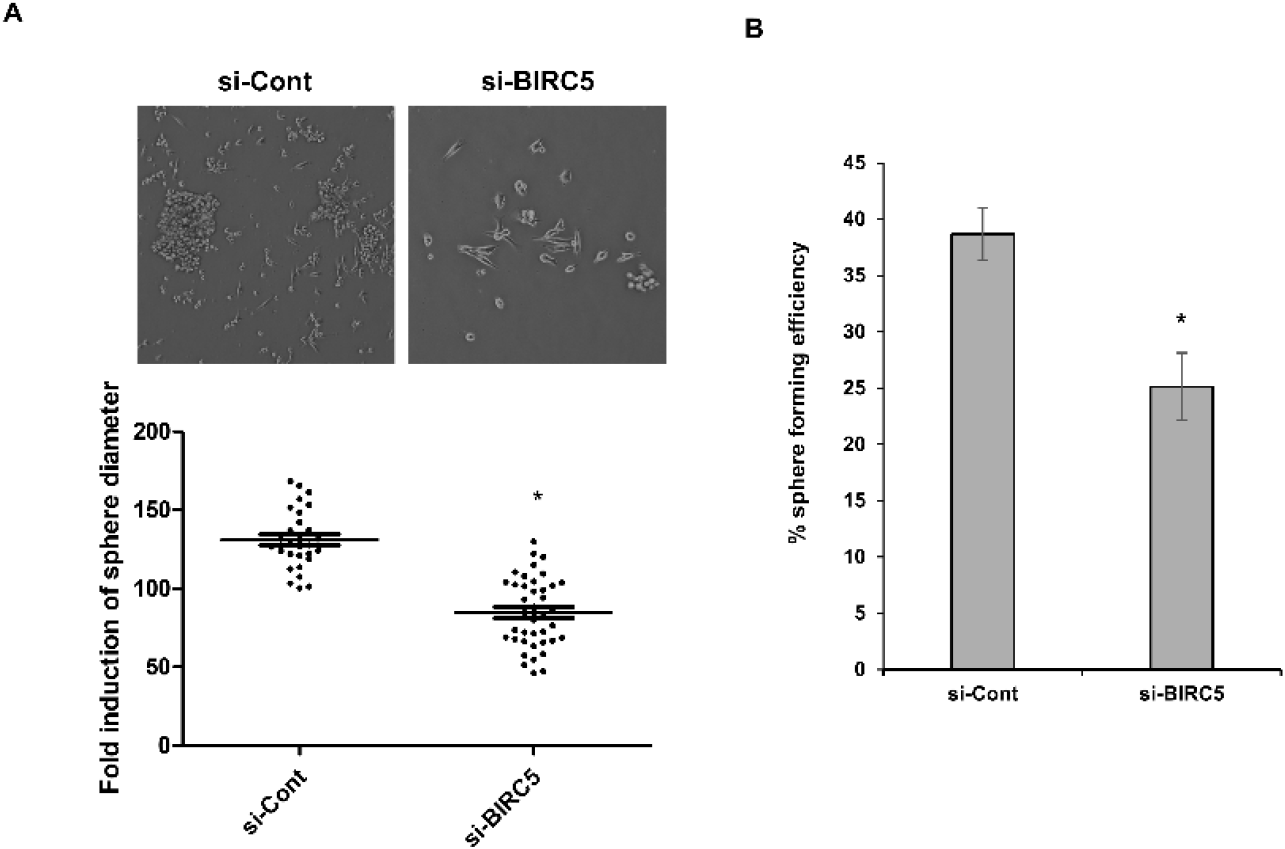
To evaluate the functional role of BIRC5 in CSC properties, sphere formation assays were performed. Silencing of BIRC5 significantly reduced both sphere diameter and sphere forming efficiency compared with control cells (Fig. 4A). To further assess self-renewal capacity at the single cell level, a single cell assay was performed. BIRC5 knockdown significantly reduced the proportion of cells capable of forming spheres compared with control cells (Fig. 4B). These results indicate that BIRC5 contributes to the self-renewal capacity of breast cancer cells.
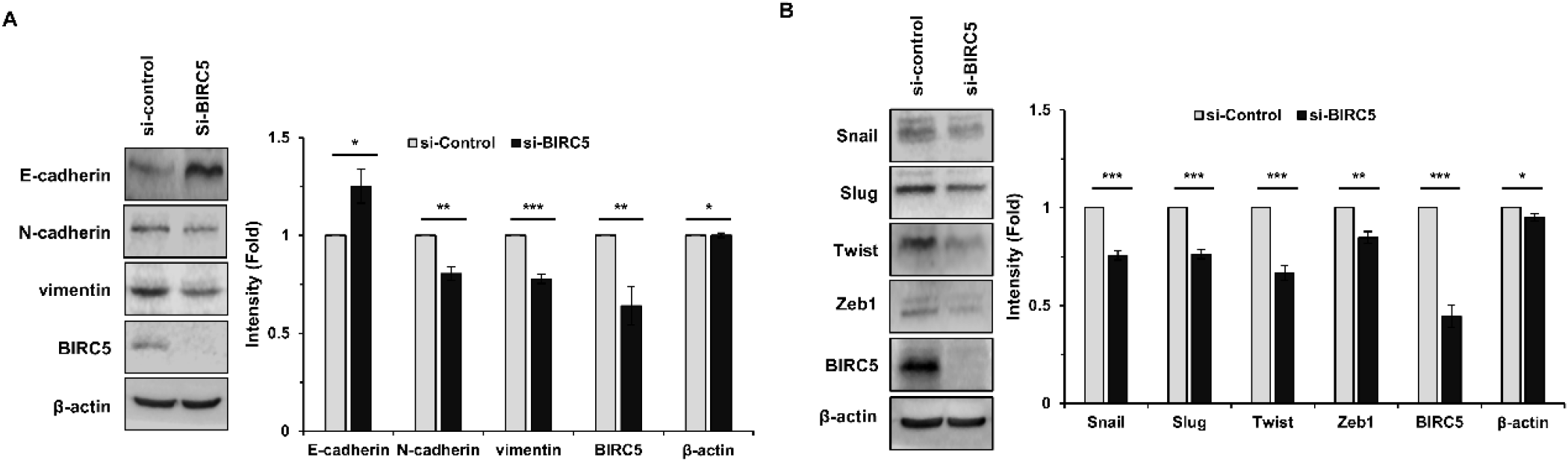
To determine whether BIRC5 is involved in EMT, the expression of EMT related proteins was examined following BIRC5 knockdown. Western blot analysis showed that silencing of BIRC5 increased the expression of the epithelial marker E-cadherin, while decreasing the expression of mesenchymal markers, including N-cadherin and vimentin (Fig. 5A). In addition, the expression of EMT associated transcription factors, such as Snail, Slug, Twist, and Zeb1, was reduced in BIRC5 silenced cells (Fig. 5B). These findings suggest that BIRC5 is associated with EMT related molecular changes.
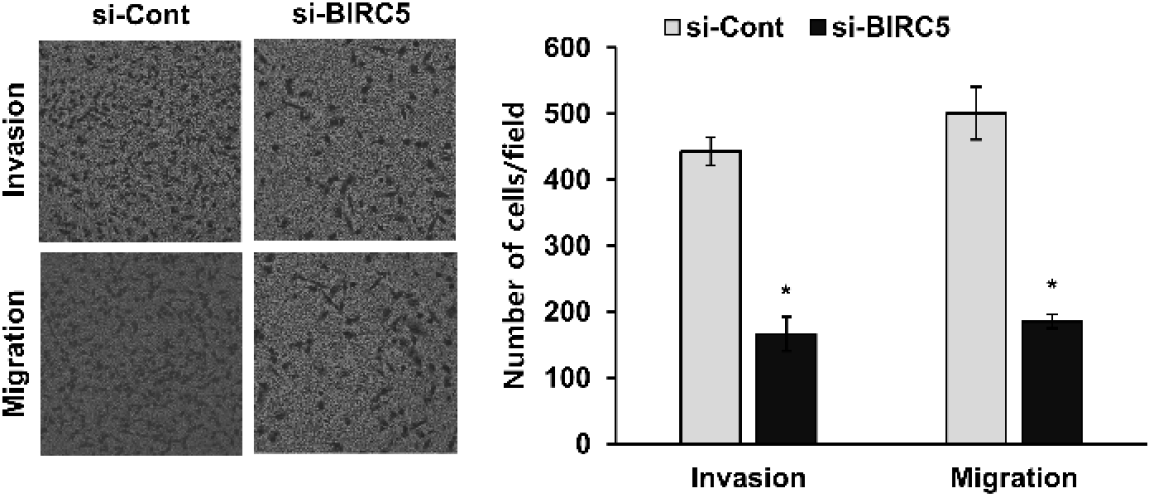
To further assess the role of BIRC5 in metastatic potential, migration and invasion assays were performed. Silencing of BIRC5 significantly reduced both cell migration and invasion compared with control cells (Fig. 6). These results indicate that BIRC5 contributes to the migratory and invasive abilities of breast cancer cells (Fig. 7).
DISCUSSION
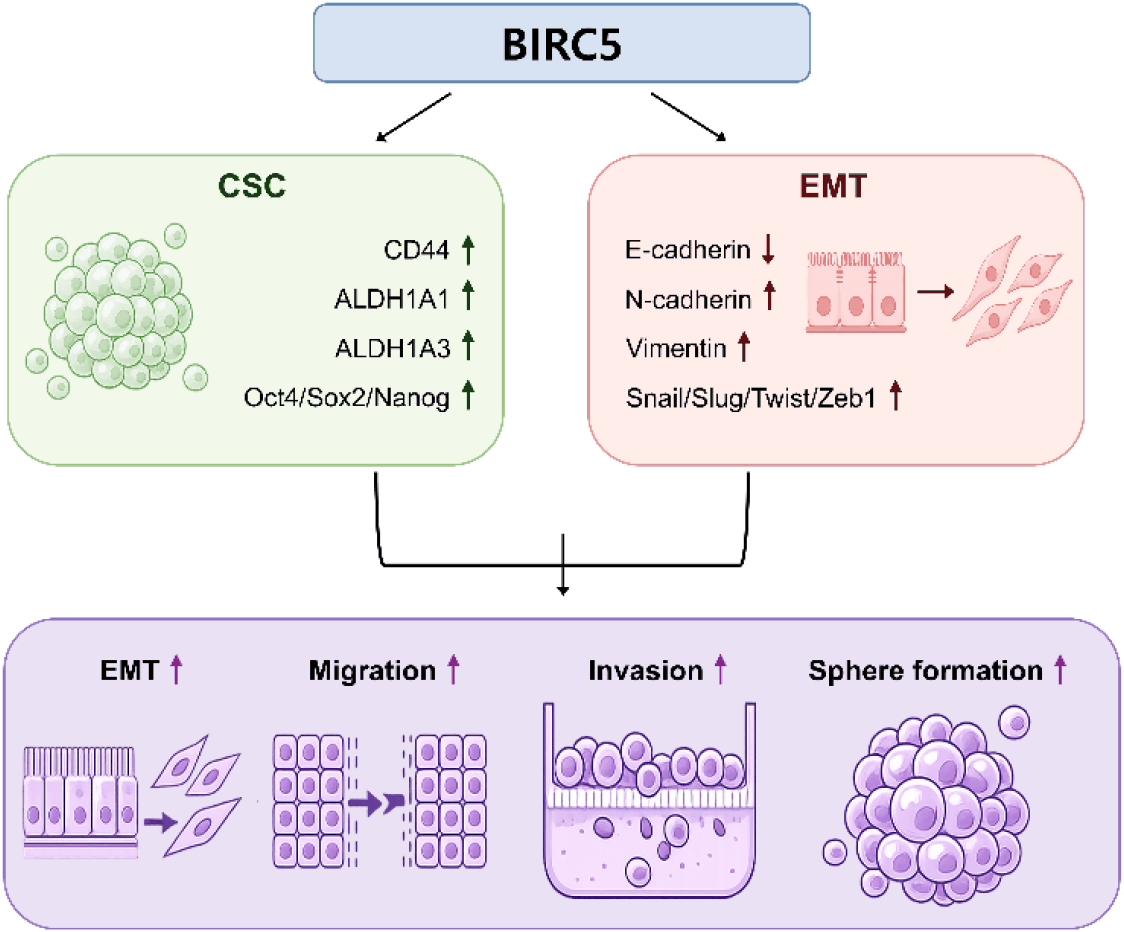
In the present study, we investigated the role of BIRC5 in the regulation of CSC properties, EMT, and metastatic potential in human breast cancer cells. Our results showed that BIRC5 expression is elevated in breast cancer and is associated with poor patient prognosis, consistent with previous reports (Altieri, 2008; Jaiswal et al., 2015). In addition, BIRC5 expression was increased under sphere forming conditions, and its silencing reduced CSC associated marker expression, sphere forming ability, EMT related protein expression, as well as migration and invasion capacities. These findings suggest that BIRC5 is involved in multiple processes associated with breast cancer progression.
CSCs are known to play a critical role in tumor initiation, progression, and therapeutic resistance due to their self-renewal capacity (Al-Hajj et al., 2003; Dean et al., 2005). In this study, sphere forming conditions increased the expression of established CSC markers, including CD44, ALDH1A1, and ALDH1A3, along with BIRC5 expression. Furthermore, knockdown of BIRC5 reduced the expression of these markers as well as pluripotency associated transcription factors such as Sox2, Oct4, and Nanog. Consistently, suppression of BIRC5 significantly decreased sphere-forming ability, indicating a functional role of BIRC5 in maintaining CSC like properties. These results are in agreement with previous studies demonstrating the association between stemness and tumor progression (Dean et al., 2005; Zhang et al., 2023).
EMT is a critical biological process that enhances the migratory and invasive abilities of cancer cells and is closely linked to metastasis (Thiery et al., 2009; Nieto et al., 2016). In the present study, silencing of BIRC5 resulted in increased expression of the epithelial marker E-cadherin and decreased expression of mesenchymal markers, including N-cadherin and vimentin. In addition, EMT associated transcription factors such as Snail, Slug, Twist, and Zeb1 were downregulated following BIRC5 knockdown. These findings are consistent with previous studies describing EMT as a key mechanism in cancer progression (Mani et al., 2008; Thiery et al., 2009).
Importantly, functional assays demonstrated that BIRC5 knockdown significantly reduced both migration and invasion abilities. In line with these findings, EMT suppression observed upon BIRC5 silencing may contribute to the reduced migratory and invasive capacities. Given that EMT and CSC properties are closely associated with metastatic progression, our results support the possibility that BIRC5 contributes to breast cancer metastasis by regulating both stemness and EMT related processes (Mani et al., 2008).
BIRC5, also known as survivin, has been widely studied for its role in inhibiting apoptosis and promoting cell proliferation (Altieri, 2008; Mani et al., 2008). Elevated expression of BIRC5 has been reported in various cancers and is often associated with poor clinical outcomes (Li et al., 1998; Mani et al., 2008). While previous studies have primarily focused on its role in cell survival, emerging evidence suggests that BIRC5 may also be involved in tumor progression beyond apoptosis regulation. Our findings extend these observations by demonstrating that BIRC5 is associated with CSC-like characteristics, EMT, and metastatic potential in breast cancer cells.
Despite these findings, several limitations should be considered. First, this study was performed using a single breast cancer cell line. To confirm these results, further validation using additional cell lines and in vivo models will be necessary. Second, the molecular mechanisms underlying BIRC5 mediated regulation of CSC properties and EMT were not investigated in detail. Future studies are required to elucidate the signaling pathways involved.
In conclusion, the present study demonstrates that BIRC5 is associated with the regulation of CSC properties, EMT, and migration/invasion abilities in breast cancer cells. These findings suggest that BIRC5 may contribute to breast cancer progression and could represent a potential target for therapeutic intervention.