INTRODUCTION
Manganese (Mn2+) is a trace metal and is essential element that is required for normal mammalian physiology. Several enzyme systems have been reported to interact with or depend on Mn2+ for their optimal catalytic or regulatory function (Gunter et al., 2006). On the other hand, excessive exposure to Mn2+ seems to cause serious neurotoxicity, immunotoxicity and developmental toxicity, particularly in male (Michalke et al., 2007).
Concerning the reproductive toxicity of Mn2+, studies have been focused on the mammalian testicular dysfunction. Chronic exposure to Mn3O4 (1,050 ppm) retarded the sexual development shown significantly smaller testis, seminal vesicle, and preputial gland weights in mice (Gray & Laskey, 1980; Webster & Valois, 1987). In human, occupational exposure to Mn2+ decreased libido and impotency (Emara et al., 1971; Mena et al., 1967), and may result in lowered sperm count and semen quality (Hjollund et al., 1998).
So far, however, research on the effect of Mn2+ exposure on female reproductive physiology has been conducted poorly. The present study aimed to evaluate the effect of 0, 1.0, 3.3, and 10 mg/kg/day doses of MnCl2 on the reproductive organs in the immature female rats.
MATERIALS & METHODS
Timely pregnant Sprague-Dawley rats were obtained from DBL (Chungcheongbuk-do, Korea) and reared in sangmyung university animal facility under conditions of 12-h light/dark cycle (lights on at 07:00 h) and constant temperature of 22±1 °C. During pregnancy and lactation, the mothers had free access to normal chow and tap water. All procedures used were approved by the Animal Care and Use Committee at Sangmyung University.
The day after weaning (postnatal day 22, PND 22), female dams were randomly assigned to the following exposure groups (n=8 dams/group) from PND 22 until PND 35: (1) oral administration of 0.9% Saline (300 μℓ /100 g BW/day, Control group), (2) oral administration of 1.0 mg/kg BW/day MnCl2 ∙ 4H2O (Sigma-Aldrich), (3) oral administration of 3.3 mg/kg BW/day MnCl2 ∙ 4H2O, and (4) oral administration of 10 mg/kg BW/day MnCl2 ∙ 4H2O. Both saline and Mn2+ solution were supplied daily for 2 weeks. At PND 35, animals were sacrificed and the tissues(ovary, uterus, oviduct, adrenal, and kidney) were immediately removed and weighed at 1,800 hour.
Uterine tissue specimens were fixed 4% paraformaldehyde then were serially dehydrated in graded ethanol and xylene. Specimens were paraffin embedded and sectioned at 5 μm thickness. Sections were stained with Hematoxylin-Eosin (H & E) stain and examined under light microscope.
The trunk blood samples were collected and centrifuged at 3,000×g for 15 min. The measurements of the serum FSH and LH were carried out according to the commercial instructions for the specific ELISA kits (USCN, China). The sensitivities of the FSH and LH assay were 0.55 ng/ml and 122.5 pg/ml, respectively. The intra-assay and inter-assay coefficients of variation were <10% and <12% for both hormones, respectively.
All values are expressed as the means (±S.E.). Differences between control and treatment groups were analysed by Student’s t-test. P values less than 0.05 were considered significant. The IBM PC programs INSTAT and PRISM 3.0 (GraphPad, San Diego, CA, USA) were used to calculate and plot the results.
RESULTS
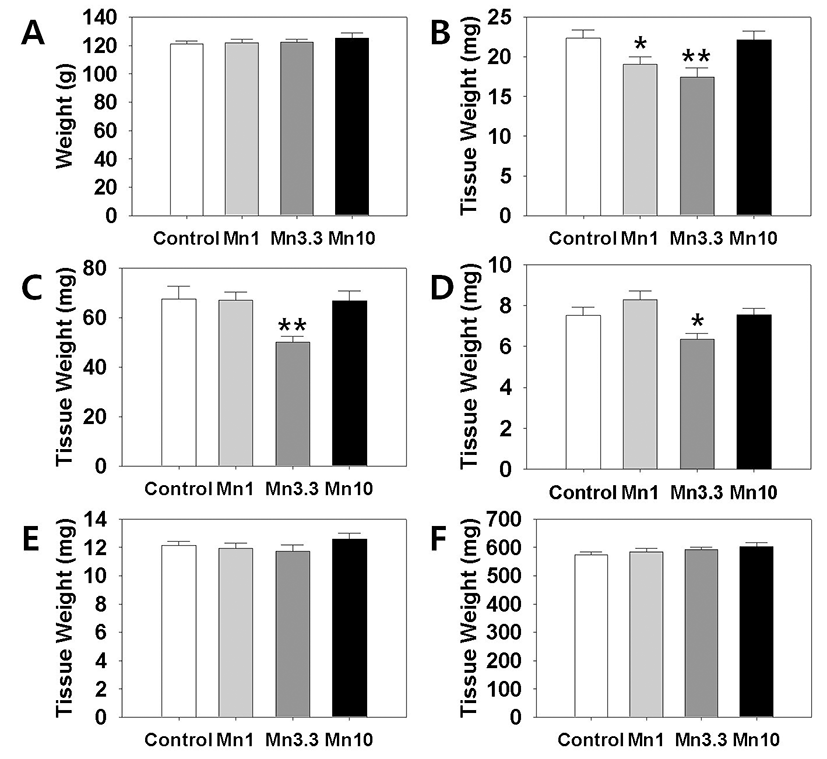
In order to evaluate a potential effect of Mn2+ on female reproductive organs, we measured the tissues weights of ovary, uterus and oviduct. Kidney and adrenal weight served as non-reproductive tissues. After 2 weeks of administration, body weights of the all Mn2+ exposure group animals were not significantly different from those of control group animals (Fig. 1, A). However, ovarian tissue weights (Fig. 1, B). in 1 mg MnCl2 dose group (19.06±0.98 mg, p<0.05) and 3.3 mg MnCl2 dose group (17.47±1.16 mg, p<0.01) were significantly lower than those of control animals (22.39±0.97 mg). Uterine tissue weights (Fig. 1, C). of 3.3 mg dose MnCl2 groups were significantly lower than those of control animals (Control : 3.3 mg dose MnCl2 = 67.52±5.15 : 50.04±2.41 mg, p<0.01), while the 1 mg MnCl2 dose (Control : 1 mg dose MnCl2=67.52±5.15 : 67.11±3.28 mg) and 10 mg MnCl2 dose (Control : 10 mg dose MnCl2 = 67.52±5.15 : 66.70±4.03 mg) failed to induce any change in uterine weight. Similarly, only 3.3 mg MnCl2 dose could induce the significant decrease in the oviduct weight (Fig. 1, D). compared to the control group (Control : 3.3 mg dose MnCl2 = 7.52±0.40 : 6.35±0.29 mg, p<0.05). Non-reproductive tissues such as adrenal (Control : 1 mg : 3.3 mg : 10 mg dose MnCl2 = 12.13±0.28 : 11.93±0.38 : 11.75±0.42 : 12.59±0.43 mg) and kidney (Control : 1 mg : 3.3 mg : 10 mg dose MnCl2 = 574.74±8.99 : 584.56±12.38 : 592.42±7.82 : 603.67±14.06 mg) failed to respond to all doses of MnCl2 exposure (Fig. 1, E & F, respectively).
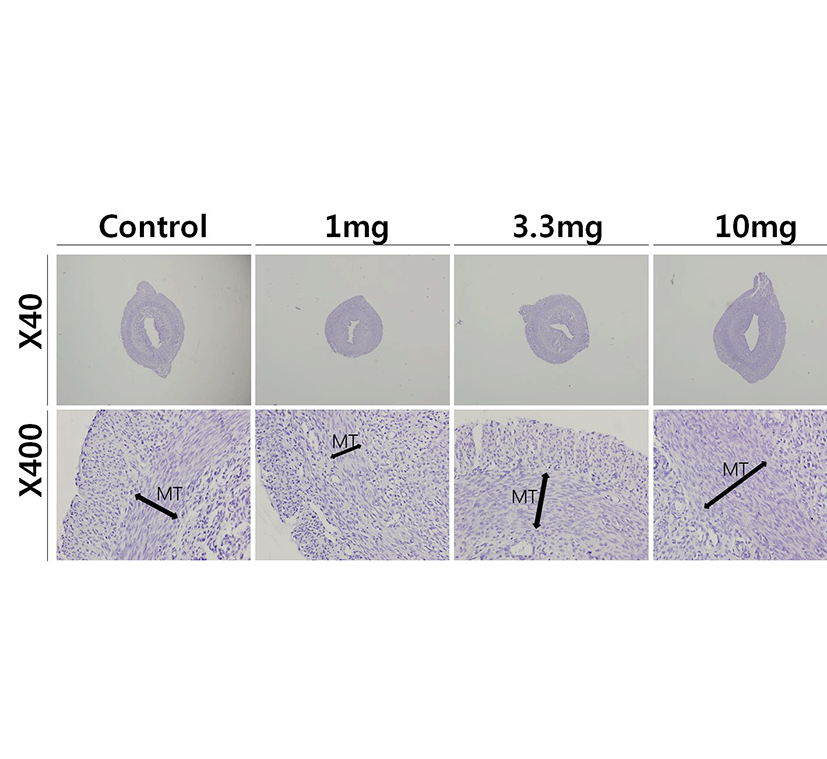
To access the histological changes in MnCl2 exposured uteri, standard paraffin section and hematoxylin-eosin staining method were employed. 3.3 mg and 10 mg MnCl2 dose groups shown the thickend myometrial layer when compared to control (Fig. 2, Mn3.3 & Mn10). In contrast, 1 mg MnCl2 exposure reduced the thickness of myometrium (Fig. 2, Mn1).
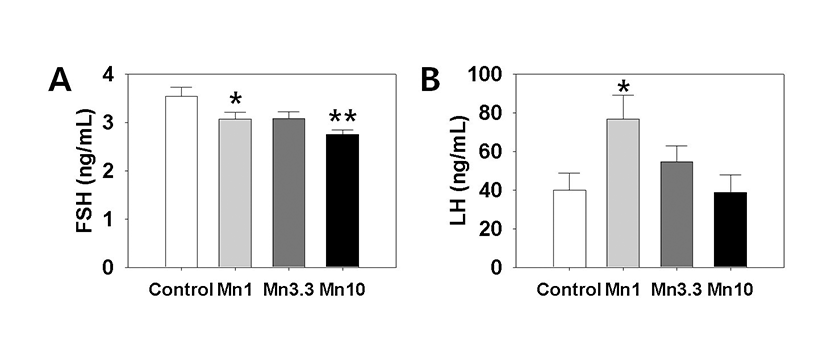
Serum LH and FSH levels were measured using specific ELISA kits. Fig. 3 (A) shows that the secretion of FSH was significantly decreased by treatment with 1 mg or 3.3 mg MnCl2 (Control:1 mg dose MnCl2 = 3.54±0.18 : 3.07±0.14 ng/ml, p<0.05; Control : 10 mg dose MnCl2 = 3.54±0.18:2.75±0.10 ng/ml, p<0.01). Serum LH level was significantly elevated by 1 mg MnCl2 exposure (Control : 1 mg dose MnCl2 = 39.85±8.93:76.71±12.36 ng/ml, p<0.05, Fig. 3, B). Higher doses of MnCl2 exposure failed to change the serum LH levels.
DISCUSSION
In the present study we demonstrated that the Mn2+ exposure could change the weights of reproductive tissues in immature female rats. Although we failed to find the advance or delay of puberty onset in the Mn2+ exposured animals (data not shown), the potential reproductive toxicity of this metal in immature female rats cannot be ruled out. Mn2+ exposure, Indeed, not only affected the proliferative activity in myometrial layer, but induced significant changes in the serum FSH and LH levels.
More defined doses and exposure periods will be helpful to verify the physiological relevance of Mn2+ exposure in female reproduction.
Occupational exposure to Mn2+ could be occurred often at the workplaces such are mines and dried battery factories (Emara et al., 1971; Mena et al., 1967). In this respect, most studies on the toxicological effects of Mn2+ have been focused on the men. Furthermore, a relatively small portion of the studies dealt with reproductive toxicity of Mn2+ exposure using male animal models. Mn2+ exposure for 2 and 4 h inhibited rat primary Leydig cell steroidogenesis by decreasing StAR protein expression while 24 and 48 h exposure of MnCl2 caused adverse effects on both StAR protein and P450scc and 3b-HSD enzyme activity to reduce steroidogenesis (Cheng et al., 2003).
Pine et al. (2005) reported that Mn2+ administered acutely into the third ventricle shown dose-dependently to stimulate LH release in prepubertal female rat, and this effect was due to a Mn2+-induced stimulation of GnRH. The authors demonstrated that Mn2+ can stimulate specific puberty-related hormones and suggested that it may facilitate the normal onset of puberty. According to them, Mn2+ may contribute to precocious puberty if an individual is exposed to elevated levels of Mn2+ too early in developmental process. This hypothesis was verified by same research group; Lee et al. (2006) demonstrated that Mn2+ is a direct stimulator of prepubertal GnRH/LH secretion and may facilitate the normal onset of male puberty. Their data that Mn2+ can cause GnRH release in adult males, suggested this action should be considered in relation to age, gender, as well as mechanistic and functional differences between adult and immature animals.
In the present study we failed to confirm the advanced puberty onset in Mn2+ exposed female rats (up to 10 mg/kg BW, data not shown). Pine et al. (2005) shown that same dose of Mn2+ exposure initiated a moderate but significant advancement in age at vaginal opening (VO) in terms of days (1.5 days). The only difference between the two studies was timing and duration of Mn2+ exposure; Our treatment regimen was PND 21-35 (14 days), and theirs was PND 12-29 (18 days). Though Mn2+ can acutely induce GnRH secretion in adult males, one should consider that additional action of Mn2+ to release GABA, a GnRH inhibitor, may ultimately contribute to suppressed reproductive function observed in adult animals following exposure to high chromic levels of Mn2+ (Prestifilippo et al., 2008). Because neurotransmitter secretory circuits in female during peripubertal period are remarkably unstable, the extent of GABA GnRH regulation, and probably more neuronal regulation, may vary responding to the minor difference. Studies indicate the more specific action mechanism of Mn2+ within the hypothalamus; Mn2+ activates soluble guanylate cyclase (sGC) directly and/or as a cofactor with available nitric oxide (NO), hence generating cGMP and resulting in prepubertal GnRH release (Lee et al., 2007). More recently, Mn2+, through the upregulation of IGF-1 and COX-2, may promote maturational events and glialneuronal communications facilitating the increased neurosecretory activity, including that of GnRH, resulting in precocious pubertal development (Hiney et al., 2011).
Special emphasis should be made on the action of Mn2+ in brain. Inhalation of the mixture of MnCl2 and Mn(OAc)3 for 5 months developed movement abnormalities, significant loss of substantia nigra compacta (SNc) dopaminergic neurons; these symptoms similar to those observed in Parkinson disease (PD) (Ordoñez-Librado et al., 2011). Dopamine (DA) depletion is closely related to pituitary prolactin biosynthesis. Male rats exposed to Mn2+ for 4 or 13 weeks showed a progressive and significant decrease in hypothalamic DA, whereas prolactin and Pit-1 mRNA levels increased in response to Mn2+ exposure (Kim et al., 2009). These results suggest that exposure to Mn2+ decreases hypothalamic DA and promotes the production of prolactin in the pituitary and that Pit-1 might be a regulator of DA and prolactin. Furthermore, such Mn2+ exposure induced significant increase in serum prolactin levels seemed to be highly correlated with the testis toxicity (Lee, 2009). In this context, relationship between Mn2+ exposure and DA/prolactin secretions in immature female rats will be helpful to understand the function of the metal during pubertal development.