
INTRODUCTION
In somatic cell nuclear transfer (SCNT) process, physical stresses generated by micromanipulation processes including enucleation, cell injection, fusion, and activation adversely affect the reprogramming of cloned embryos by inducing cellular stresses such as oxidative and endoplasmic reticulum (ER) stresses. (Hwang et al., 2012, 2013; Park et al., 2017; Lee et al., 2018).
Previous studies showed that ER stress and reactive oxygen species (ROS) levels of SCNT embryos were decreased by treatment of ER stress inhibitor and antioxidant during the culture period (Yoon et al., 2014). In general, ER stress inhibitor and antioxidant are treated during the maturation period of oocytes or culture period of embryos (Yoon et al., 2014; Mallol et al., 2015; Lin et al., 2015, 2016). In contrast, previous studies showed that development and quality of SCNT embryos were improved by treating ER stress inhibitors only during microinjection and activation of SCNT, inhibiting ER stress generation during SCNT, and reducing apoptosis (Park et al., 2017; Park et al., 2019). In addition, the ROS and apoptosis were reduced when antioxidant was treated during micromanipulation, fusion and activation of SCNT process (Bae et al., 2012, 2015).
An intimate correlation between ER stress and oxidative stress has been demonstrated by many physiological and pathological studies (Cullinan & Diehl, 2006; Malhotra & Kaufman, 2007; Malhotra et al., 2008; Bhandary et al., 2012; Zeeshan et al., 2016). Previous researches have demonstrated that ER stress and ROS are not only occurring simultaneously, but also inducing one another (Cullinan & Diehl, 2006; Eletto et al., 2014; Zhang et al., 2015; Zeeshan et al., 2016). In addition, PKR-like ER kinase (PERK), one of the unfolded protein response (UPR) sensors, regulates the expression of detoxifying enzymes and genes involved in protein folding by inducing the phosphorylation of the nuclear factor (erythroid-derived 2) 2 (Nrf2), the master key of the antioxidative response (Cullinan & Diehl, 2006; Bhandary et al., 2012; Zeeshan et al., 2016). Conversely, high levels of ROS have been reported to promote ER stress by damaging cellular proteins such as abnormal folding, aggregation and fragmentation, inhibiting protein production and transport, and destroying calcium homeostasis (Malhotra & Kaufman, 2007; Vannuvel et al., 2010; Sano & Reed, 2013). However, it is unclear how the correlation between ER stress and ROS affects the reprogramming and development of early embryos.
Taken together, the intimate interaction between ER stress and oxidative stress, which are generated during micromanipulation, may interactively affect the development and reprogramming of SCNT embryos. Thus, it is surmised that ER stress inhibitor and antioxidant treatments during SCNT may effectively inhibit cell damage and promote the reprogramming of SCNT embryos. In this study, we focused on the close interaction between ER stress and ROS during SCNT process. Thus, we investigated the effects of the treatment of tauroursodeoxycholic acid (TUDCA), an ER inhibitor, and vitamin C (Vit. C), an antioxidant during micromanipulation, on the ER stress and oxidative stress of SCNT embryos, and on the cell damage and development of SCNT embryos.
MATERIALS AND METHODS
All chemicals and reagents were supplied by Sigma-Aldrich Chemical Co. (St. Louis, MO, USA) unless otherwise specified. Vit. C and TUDCA (Merck, Darmstadt, Germany) stocks dissolved in phosphate buffered saline (PBS, Gibco, Grand Island, NY, USA) were stored at −20°C until use.
Porcine cumulus-oocyte complexes (COCs) were aspirated from follicles (3 to 6 mm in diameter) using 18-gauge needle 10 mL syringe. COCs were washed with Tyrode’s lactate (TL)-HEPES buffer containing 0.1% (w/v) polyvinyl alcohol (PVA), then cultured in in vitro maturation (IVM) medium for 42–44 h at 39°C, 5% CO2 in air. IVM medium was Tissue Culture Medium 199 (TCM199; Gibco) supplemented with 0.1% (w/v) PVA, 3.05 mM D-glucose, 0.91 mM Na-pyruvate, 0.57 mM cysteine, 75 μg/mL penicillin G, 50 μg/mL streptomycin, 0.01 IU/mL follicle stimulating hormone (FSH), 0.01 IU/mL luteinizing hormone (LH), and 10 ng/mL epidermal growth factor (EGF).
Porcine ear skin cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Gibco) supplemented with 10% fetal bovine serum (FBS; GenDEPOT, Katy, TX, USA) and 1% (w/v) penicillin and streptomycin (P/S; Mediatech, Manassas, VA, USA) at 39°C, 5% CO2 in air. The cells were passaged 2–3 times and frozen using DMEM containing 10% dimethyl sulfoxide (DMSO; Junsei Chemical, Tokyo, Japan) and stored in liquid nitrogen. Prior to SCNT, cells were thawed and cultured in DMEM containing 15% FBS and 1% (w/v) P/S at 39°C, 5% CO2 in air until they reached confluence to synchronize the cell cycle stage at the G0/G1 phase. Cells were trypsinized with 0.05% (w/v) trypsin-EDTA (Gibco) and washed by centrifugation (500×g, 4 min, room temperature) in Hepes-buffered TCM199 supplemented with 0.78 mM sodium bicarbonate, 0.08 mM streptomycin, 0.14 mM penicillin G, and 3 mg/mL bovine serum albumin (BSA). Donor cells were placed in a 500 μL well of the same medium for use.
The cumulus cells of maturated oocytes were eliminated by vortexing in PBS containing 0.1% (w/v) hyaluronidase and 0.1% (w/v) polyvinyl pyrrolidone (PVP) for 3 min. Enucleation of oocytes was conducted by removing the metaphase II (MII) first polar body and chromosome mass using injection pipette in the micromanipulation medium of Hepes-buffered TCM-BSA containing 5 μg/mL cytochalasin B. More than 90% of the oocytes were enucleated with this method (data not shown). A single donor cell was then injected into the perivitelline space of an enucleated oocyte.
Reconstructed oocytes were placed in Porcine zygote medium-3 (PZM-3, holding medium) for 10-30 min prior to electrofusion. Then, they were placed and aligned between two wire electrodes (1 mm apart) of an electrofusion chamber, overlaid with 0.3 M mannitol solution containing 0.1 mM MgCl2, 0.1 mM CaCl2 and 0.5 mM Hepes (Duchefa Biochemie, Haarlem, Netherlands). For electrofusion/activation, a single direct current pulse of 1.25 kV/cm was applied for 30 μsec using a BTX Electro Cell Manipulator 200 (BTX, San Diago, CA, USA). After fusion/activation, the reconstituted oocytes were placed in holding medium at 39°C, 5% CO2 in air and checked for fusion.
For the treatment groups, 100 μM TUDCA (Park et al., 2019), 100 μM Vit. C (Bae et al., 2012), or both TUDCA and Vit. C (TUD+Vit. C) were added to micromanipulation medium and holding medium prior to fusion of reconstituted oocytes. After fusion/activation, the reconstituted oocytes for the treatment group were further incubated in the PZM-3 medium containing ER stress inhibitors and/or antioxidant (TUDCA, Vit. C or TUD+Vit. C, respectively) of each concentration at 39°C, 5% CO2 in air for 3 h.
After fusion/activation (control) or treatment of ER stress and oxidative stress inhibitors, SCNT embryos were cultured in a 40 μL droplet of fresh PZM-3 medium at 39°C, 5% CO2 in air for up to 6 days. At the 1-cell stage (20 h) or blastocyst stage (6 days), the embryos were washed in PBS supplemented with 0.3% (w/v) PVP (PBS-PVP) and lysed by using 30 μL of Lysis/Binding (L/B) buffer of Dynabeads® mRNA Direct kit™ (Life Technologies, Oslo, Norway). The lysed embryo samples were stored at −70°C until use.
The blastocysts were washed with PVP-PBS, then stained with 20 μg/mL of Hoechst 33342 for 30 min. After staining, the embryos were mounted on a slide glass with Vecta-Shield (Vector Laboratories, Burlingame, CA, USA) and covered with a coverslip. The number of cells in blastocysts were counted by using a fluorescence microscopy (BX50, Olympus, Tokyo, Japan).
Poly(A) mRNA of the SCNT embryos was isolated by using Dynabeads® mRNA Direct kit™ (Life Technologies) according to the manufacturer’s protocols. The cryopreserved embryo samples were thawed and mixed with 30 μL of Dynabeads oligo (dT)25 by shaking for 8 min at room temperature to allow the hybridization of poly (A) tail of mRNA with the oligo (dT)25 on the beads. The Beads-mRNA complexes were washed twice with 100 μL of wash buffer A and B, respectively. Beads were separated from supernatant by using a DynaMag™-Spin Magnet (Invitrogen, Carlsbad, CA, USA). The poly (A) mRNA was eluted from beads by incubation with elution buffer (12.5 μL of 10 mM Tris-HCl) for 5 min at 70°C. The cDNA synthesis was performed using AccuPower® RocketScript™ Cycle RT Premix (Bioneer, Daejeon, Korea) according to the manufacturer’s protocol. Each 10 μL mRNA was used as a template. The reaction was carried out in Veriti® 96-well Thermo cycler (Applied Biosystems, Foster City, CA, USA) by 5 cycles of 4°C for 5 min, 37°C 15 sec, 50°C for 5 min, and 98°C for 5 min. The cDNA products were stored at 4°C until use.
Semi-qPCR analysis was used to detect the Xbp1 mRNA expression. The expressions of ER stress-associated genes such as binding protein (BiP), glucose-regulated protein 94 (GRP94), C/EBP homologous protein (CHOP), and activating transcription factor 4 (ATF4), and apoptotic genes such as Bcl2-associated X protein (Bax) and caspase-3, and oxidative stress-associated genes, such as Nrf2 and hemeoxygenase 1 (HO-1) were analyzed by RT-qPCR. For Semi-qPCR, the cDNA sample was reverse-transcribed using AccuPower® Taq PCR PreMix (Bioneer) according to the manufacturer’s protocol. PCR conditions were consisted of a denaturation at 95°C for 30 sec, an annealing at 58°C for 30 sec, and an extension at 72°C for 5 min. The PCR products were analyzed by UV irradiation with a Gel Doc™XR+ (Bio-rad, Berkeley, CA, USA) on 4% agarose gel (Amresco, Cleveland, OH, USA) supplemented with 0.05% ethidium bromide (Bioneer). RT-qPCR was performed using 1 μL cDNA with SYBR Green PCR Master Mix (TOPreal™ qPCR 2X PreMIX; SYBR Green with high ROX, Enzynomics, Daejeon, Korea). The amplification reaction was performed in a StepOne Plus instrument (Applied Biosystems) by 40 cycles of a denaturation at 95°C for 30 sec, an annealing at 60°C for 30 sec, and an extraction at 72°C for 30 sec. The comparative CT method (ΔΔCT method) was used for the relative mRNA quantification of each target gene. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as a housekeeping gene. Primer pairs synthesized for the primer sequences of each gene are shown in Table 1.
Xbp1, X-box binding protein 1; BiP, binding protein; GRP94, glucose-regulated protein 94; ATF4, activating transcription factor 4; CHOP, C/EBP homologous protein; Nrf2, nuclear factor 2; HO-1, hemeoxygenase 1 GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
At least 5 replicates were conducted for each statistical analysis. Data of semi-qPCR and RT-qPCR were analyzed by the general linear model (GLM) and the Duncan’s multiple range test. The developmental rate was analyzed by Chi-square test, and the blastocyst cell number was analyzed by Student t-test. All analyses were performed using the Statistical Analysis System software package (v.9.4, SAS Institute, Cary, NC, USA).
RESULTS
TUDCA and Vit. C treatment groups showed the significant reduction of Xbp1 splicing compared to the control group at the 1-cell stage (p<0.05). However, Xbp1 splicing in the combined group (TUD+Vit. C) was not different from that of the control group (Fig. 1A).
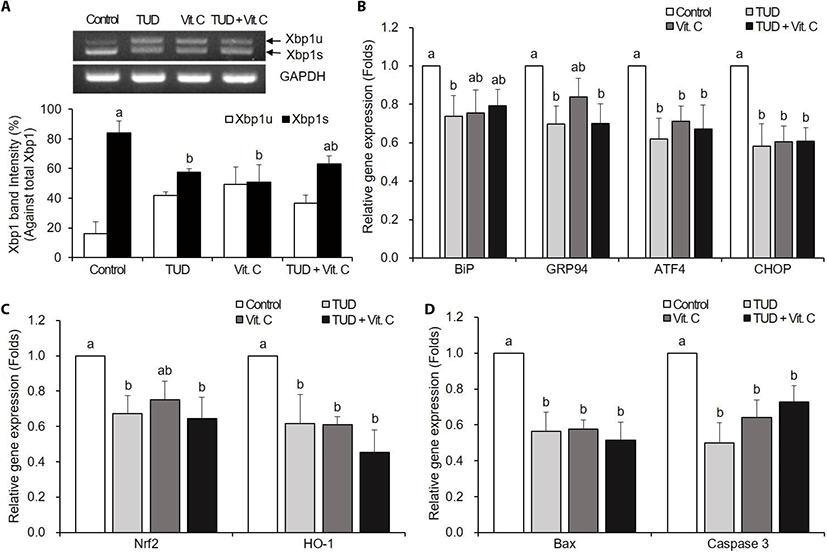
The expressions of ER stress-associated genes BiP, GRP94, ATF4, and CHOP were significantly decreased in all treatment groups compared to the control group (p<0.05), except BiP in Vit. C and TUD+Vit. C treatment groups, and GRP94 in Vit. C treatment group (Fig. 1B).
The expressions of oxidative stress-associated genes Nrf2 and HO-1 tend to decrease in all treatment groups (Fig. 1C). Nrf2 expression was significantly decreased by TUDCA and TUD+Vit. C treatments, and HO-1 expression was significantly decreased by all treatments compared to control (p<0.05).
The transcription levels of pro-apoptotic genes Bax and caspase-3 mRNAs were significantly decreased (p<0.05) in all treatment groups compared to the control group (Fig. 1D).
There were no significant differences in Xbp1 splicing among all groups (Fig. 2A).
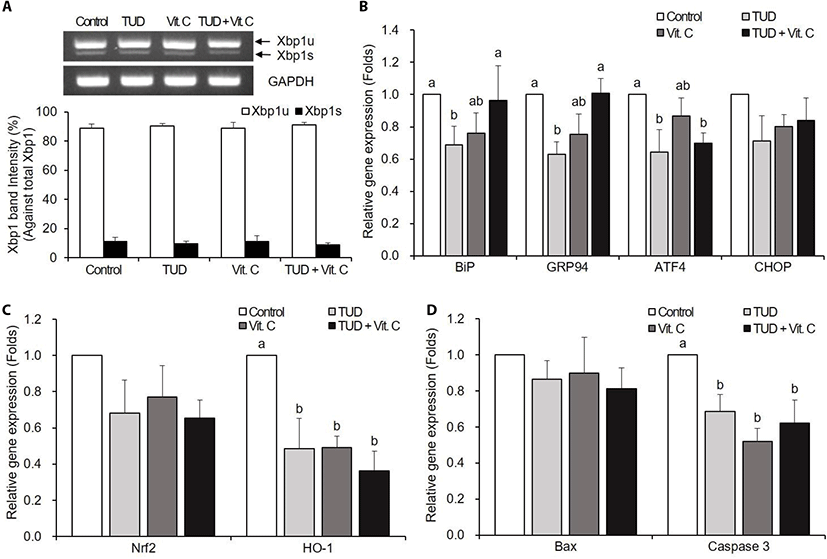
The Vit. C treatment did not reduce the expression levels of ER stress-associated genes of porcine SCNT embryos at the blastocyst stage (Fig. 2B). However, TUDCA treatment significantly reduced the expression levels of BiP, GRP94, and ATF4, and the TUD+Vit. C treatment reduced only ATF4 transcription level (p<0.05).
At the blastocyst stage, ER stress inhibitor and antioxidant treatments significantly decreased (p<0.05) the expressions of oxidative stress-associated gene HO-1, but not Nrf2 (Fig. 2C).
There were no significant differences in Bax mRNA expression compared to control (Fig. 2D). However, the transcription levels of caspase-3 mRNA significantly reduced in all treatment groups compared to the control group (p<0.05).
Blastocyst formation rate was tended to increase in all treatment groups, but significantly increased in TUDCA (24.8%, 55/222) and Vit. C (22.0%, 53/241) treatment groups compared to the control group (14.7%, 39/265, p<0.05). The mean cell number in blastocysts was significant increased in all treatment groups (59.7±4.3 to 63.5±3.3) compared to the control group (46.2±2.6, p<0.05, Table 2).
Experiments were repeated 6 times in each treatment group.
TUD, 100 μM TUDCA (tauroursodeoxycholic acid); Vit. C, 100 μM vitamin C.
DISCUSSION
The signal pathway of UPR sensor proteins activates ATF4, CHOP, Xbp-1, and activating transcription factor 6 (ATF6), which promote the expression of ER-associated degradation (ERAD)-related genes BiP, GRP94, and ER chaperone genes to restoring the homeostasis of ER (Ron & Walter, 2007; Senft & Ronai, 2015). When oxidative stress occurs, Nrf2 is activated through phosphorylation. Subsequently, Nrf2 transfer to the nucleus and binds to the antioxidant elements (ARE) to regulate the expression of many antioxidant genes including superoxide dismutase 1 (SOD1), catalase and HO-1 (Cullinan & Diehl, 2006; Nguyen et al., 2009; Zhang et al., 2015).
TUDCA is a chemical chaperone known as an ER stress inhibitor that assists protein folding and suppresses ER stress (Xie et al., 2002; Lee et al., 2010). Several studies have shown that ER stress is inhibited when TUDCA is treated to embryo or cell in culture period (Zhang et al., 2012a,b; Yoon et al., 2014; Lin et al., 2015, 2016, Mochizuki et al., 2018). In addition, our previous study showed that when TUDCA is treated during the micromanipulation and activation processes of the SCNT embryo, Xbp1s and ER stress-associated genes are significantly reduced, thereby increasing the development and quality of SCNT embryos (Park et al., 2019). This study substantiates our previous result. Interestingly, the TUDCA treatment reduced not only ER stress but also oxidative stress-associated gene transcription levels at the 1-cell stage in this study. These results suggest that TUDCA restores ER function by stabilizing protein folding and induces the decrease in ROS through uncertain mechanism.
Several studies suggested that ROS induce ER stress through regulation with UPR signaling (Cullinan & Diehl, 2006; Eletto et al., 2014; Zhang et al., 2015). Thus, the reduction of ROS through antioxidant treatment may lead to the inhibition of ER stress. In previous study, antioxidant treatment resulted in the suppression of ER stress and increased secretion of normal protein (Malhotra et al., 2008). Also, similar results were obtained when the antioxidant N-acetylcysteine was administered to mouse hepatocytes (Lin et al., 2013). Yoon et al. (2014) also reported that ER stress levels of SCNT embryos decreased by treatment of antioxidant during the culture period. These results corroborate our present result that antioxidant treatment during SCNT process reduced ER stress by decreasing Xbp1 splicing and ER stress-associated gene transcription levels in the 1-cell stage.
There was no significant decrease in the splicing of Xbp1 in combination with ER stress inhibitor and antioxidant. Previous study showed that the combination of ER stress inhibitor TUDCA and salubrinal did not significantly decrease the Xbp1s and ER stress-associated gene transcription levels, suggesting that simultaneous treatment of both agents would have had an excessive effect on ER (Park et al., 2019). From this point of view, a similar tendency would have been observed when the ER stress inhibitor and antioxidant were simultaneously treated. Though, the expression levels of ER stress-associated genes (BiP, GRP94, ATF4, and CHOP) and oxidative stress-associated genes (Nrf2 and HO-1) were significantly reduced at the 1-cell stage, thus combined treatment may also have an effect on reducing ER stress and ROS.
In the 1-cell stage, apoptosis was inhibited by the treatments of ER stress inhibitor and/or antioxidant. When UPR is activated by ER stress, the activity of CHOP protein and IRE1-mediated apoptosis signal-regulating kinase 1 (ASK1) / c-Jun N-terminal kinase (JNK) causes excessive Ca2+ release in the ER, resulting in increased mitochondrial Ca2+ concentration (Nishitoh et al., 2002; Zeng et al., 2015). Increased mitochondrial Ca2+ concentration activates caspase-3 by the intrinsic pathway and increases apoptosis by increasing the expression of pro-apoptotic protein Bax (Song, 2012; Sano & Reed, 2013). In particular, CHOP is involved in transcription of genes such as death receptor 5 (Dr5), which are involved in cell death, and directly affect apoptosis (Yamaguchi & Wang, 2004). On the other hand, since antioxidant treatment can suppress apoptosis by decreasing ROS, it cannot be asserted that the decrease in apoptosis in this study is the result of the suppression of ER stress completely (Salganik, 2001; Bae et al., 2012, 2015).
ER stress inhibitor and antioxidant were not effective in preventing ER stress but affected only some of the oxidative stress-related genes and apoptotic genes in the blastocyst stage. Whereas, in previous studies, there was a difference in gene expression in the blastocyst stage when TUDCA was treated for culture period (Zhang et al., 2012a,b; Yoon et al., 2014). In this study, however, the ER stress was suppressed during only micromanipulation and activation, and the short term of initial culture rather than the whole culture period. The ER stress inhibitor treatment during the culture period would have continued to affect the expression of the genes of blastocyst. Although, when the antioxidant and / or ER stress inhibitor were treated during SCNT process in this study, there was no significant effect in ER stress and apoptosis in the blastocyst stage, the blastocyst development and cell number in blastocyst were improved.
In conclusion, ER stress and ROS are closely correlated to each other and co-regulate. Furthermore, it was illustrated that the treatment of antioxidant and / or ER stress inhibitor during micromanipulation of SCNT process can reduce the oxidative stress as well as the ER stress and the apoptosis of SCNT embryos in the early embryonic stage, and consequently promote developmental competency and the reprogramming of SCNT embryos.