
INTRODUCTION
Proper maturation of spermatozoa released from the testis is chiefly occurred in the epididymis and absolutely required for successful male reproduction. Depending on morphological and functional characteristics, the epididymis is divided into initial segment (IS), caput epididymis, corpus epididymis, and cauda epididymis (Cosentino & Cockett, 1986). The epithelium of the IS is composed of several cell types, including principal, basal, and halo cells, which are frequently found throughout the entire epididymis (Robaire & Hermo, 1988). However, the epithelial layer also has narrow and apical cells that are exclusively present in the IS (Robaire & Hermo, 1988). Several researches have demonstrated that the luminal composition of the IS is distinguishable from the other parts of the epididymis (reviews in Robaire & Hermo, 1988). In addition, distinct epithelial localization of enzymatic molecules indicates that the function of the IS would be different from the rest of the epididymal region (Adamali & Hermo, 1996). Together, these observations imply that functional regulation of the IS is likely controlled by a different manner from other epididymal parts.
Precisely regulated coordination among cells is absolutely required to maintain functions of a tissue. Specially, direct interaction between neighboring cells is important in the functional regulation of mammalian tissues, including the male reproductive organs (Pointies et al., 2005). There are three different types of direct cell-cell interactions, such as cadherin-based adhesion, occluding-based attachment, and connexin (Cx)-based gap junction (Pointies et al., 2005). Of these, only Cx-based gap junction allows direct exchange of intracellular exchange of signal molecules between cytoplasm of adjacent cells (Goodenough et al., 1996). The gap junction is a complex of two homo- or heteromeric hemichannels, which each hemichannel is consisted of six Cxs (Cyr, 2011). More than 20 Cx isoforms in mammals have been identified up to date (Meşe et al., 2007). Several researches have demonstrated the presence of Cxs in the male reproductive tract (Cyr, 2011; Lee, 2013; Seo et al., 2010). Segment-specific expression of Cxs in the epididymis during postnatal development has been reported (Dufresne et al., 2003). Our previous study showed the expression of 8 Cxs, including Cx30.3, 31, 31.1, 32, 37, 40, 43, and 45, in the IS during postnatal period (Seo et al., 2010). Expressional levels of some Cxs are generally high at puberty and adulthood, while others have shown the highest expression at prepubertal age (Seo et al., 2010). Expression of Cx32 and 37 in the IS during postnatal period is fluctuated (Seo et al., 2010). These observations indicate the complexity of expressional regulation of Cx genes in the IS. Less attention has been paid on the regulation of Cx gene expression in the IS. St-Pierre et al. (2003) have shown that induction of hypothyroidism during neonatal period results in a decrease of Cx43 expression in the IS of adult rat, implying hormonal regulation of the expression of Cx genes in the IS. Another study demonstrates a possibility of expressional regulation of Cx 43 by epidermal growth factor in human cauda epididymis (Dubé et al., 2012). However, more detailed examination has not been attempted to reveal expressional regulation of Cx genes in the IS.
It is well known that the epididymis is steroid hormoneresponsive tissue. There is no doubt that the function and structure of epididymis are chiefly under the regulation of androgens (Robaire & Hamzeh, 2011). In addition, there are growing numbers of evidence of demonstrating the effect of estrogens on the regulation of epididymal functions (Joseph et al., 2011). Such effects of androgens and estrogens on the epididymis are exerted by modulating the expression of various genes associated with structural and functional regulation of the epididymis (Joseph et al., 2011; Robaire & Hamzeh, 2011). Yet, the effect of steroid hormones on expression of Cx genes in the IS has not been thoroughly studied. As mentioned earlier, our previous research shows the existence and differential expressional patterns of several Cx isoforms in the IS during postnatal development (Seo et al., 2010). Thus, in the present study, we attempted to show if neonatal treatment of flutamide, an anti-androgen, or estradiol benzoate, an estrogen agonist, gives an influence on the expression of Cx genes in the IS of adult rat.
MATERIALS AND METHODS
Five pregnant female Spragure Dawley rats were purchased from Samtako (OSan, Korea) and individually caged upon the arrival. Food and drinking water were free access to food and drinking water for the entire experimental period. Each pregnant female rat was randomly assigned into one of five different experimental groups, control, low-dose EB-treated (EB-L), high-dose EB-treated (EB-H), low-dose Flu-treated (Flu-L), or high-dose Flu-treated (Flu-H) group. At the end of the term, 5 to 8 male pups were delivered from each female rat.
The powder of EB and Flu purchased from Tokyo Chemical Industry Co. (Tokyo, Japan) was completely dissolved in 100% EtOH at room temperature, and then diluted in sesame oil. When the pups became 7 days of postnatal age, each pup was weighted and subcutaneously injected by EB at the final concentration of either 0.015 or 1.5 μg/kg body weight (BW) for EB-L or EB-H, respectively, or Flu at the final concentration of either 500 μg or 50 mg/kg BW for Flu-L or Flu-H, respectively. Control animals were treated by sesame oil of same amount.
Once sexually mature at 4 months of age, the animals were anesthetized by CO2 stunning and the reproductive tract was exposed through an incision on lower abdomen. The reproductive tract was placed in cold PBS, and the epididymis was carefully dissected out from the rest parts of the reproductive tract. The IS was separated from the epididymis and washed a couple of times in fresh cold PBS before freezing in liquid nitrogen. The IS was stored in –80°C until used for total RNA isolation. In total, 31 male rats were used for the present research, including control (n=5), EB-L (n=7), EB-H (n=7), Flu-L (n=6), and Flu-H (n=6).
To isolate total RNA from the tissues, frozen IS was first homogenized in easy-Blue total RNA extraction solution (iNtRON Biotech, Sungnam, S.Korea) with a polytron homogenizer (Fisher Scientific, Pittsburgh, PA). Then, using phenol-chloroform extraction method, a pellet of total RNA was collected and resuspended in DEPC-treated dH2O. Qualitative evaluation of isolated total RNA was carried out with gel electrophoresis, and the amount of total RNA was acquired by quantitative analysis with an UV spectrophotometer (Eppendorf, New York, USA). The total RNAs were stored in –80°C until utilized for cDNA generation.
The first strand of cDNA was generated from 1 mg of total RNA, using oligo-dT primer and ImProm-II™ reverse transcription system (Promega, Madison, USA). The reverse transcription (RT) reaction was carried out at 25°C for 5 min, 42°C for 1 hr and 30 min, and 70°C for 15 min. The cDNA was directly used for quantitative real-time PCR to determine expressional levels of connexin (Cx) isoforms in the IS of experimental animals. Table 1 presents information of oligonucleotide primers utilized for real-time PCR analysis in the present study.
The PCR cocktail was prepared with 1 mL of cDNA, 10 pmol of primer set, 10 μL of master mixture (Finnzymes, Espoo, Finland), and water to make a final volume of 20 μL. The PCR was carried out by cycles of denaturation at 95°C for 30 sec, annealing at Tm for 30 sec, and extension at 72°C for 30 sec. At the end of each PCR, an additional extension step at 72°C for 10 min was included. Cyclophilin A (Ppia) was used as a PCR internal quantitative control. Expected sizes of PCR products were confirmed by 1.2% agarose gel electrophoresis.
The RT reaction and PCR were independently quadruplicated to get a mean and a standard error for each experimental group. Experimental results are present in relative ratios of expression between Ppia and Cx isoform. Statistical comparisons of gene expression levels among experimental groups of each treatment were carried out by one-way ANOVA, followed by a post-hoc analysis, Duncan’test. When P value was lower than 0.05, the result was considered as statistically significant.
RESULTS
Expression of Cx30.3 in the IS at the adult was significantly increased by EB treatment at 1 week of age, even though there was no difference on Cx30.3 expressional level between low and high dose treatment of EB (Fig. 1a). A treatment of low-dose Flu caused no significant change in Cx30.3 mRNA level in the IS, even though a high-dose Flu treatment at 1 week of age resulted in a significant elevation of Cx30.3 expression in the adult IS (Fig. 1a). Expressional level of Cx31 in the adult IS was not affected by low-dose EB treatment at 1 week of age (Fig. 1b). But, a treatment of high-dose EB led to a significant decrease of Cx31 transcript level (Fig. 1b). In contrast with Cx31, a treatment of low-dose Flu caused a significant decrease of Cx31 expression, while there was no change on Cx31 mRNA level by high-dose Flu treatment (Fig. 1b).
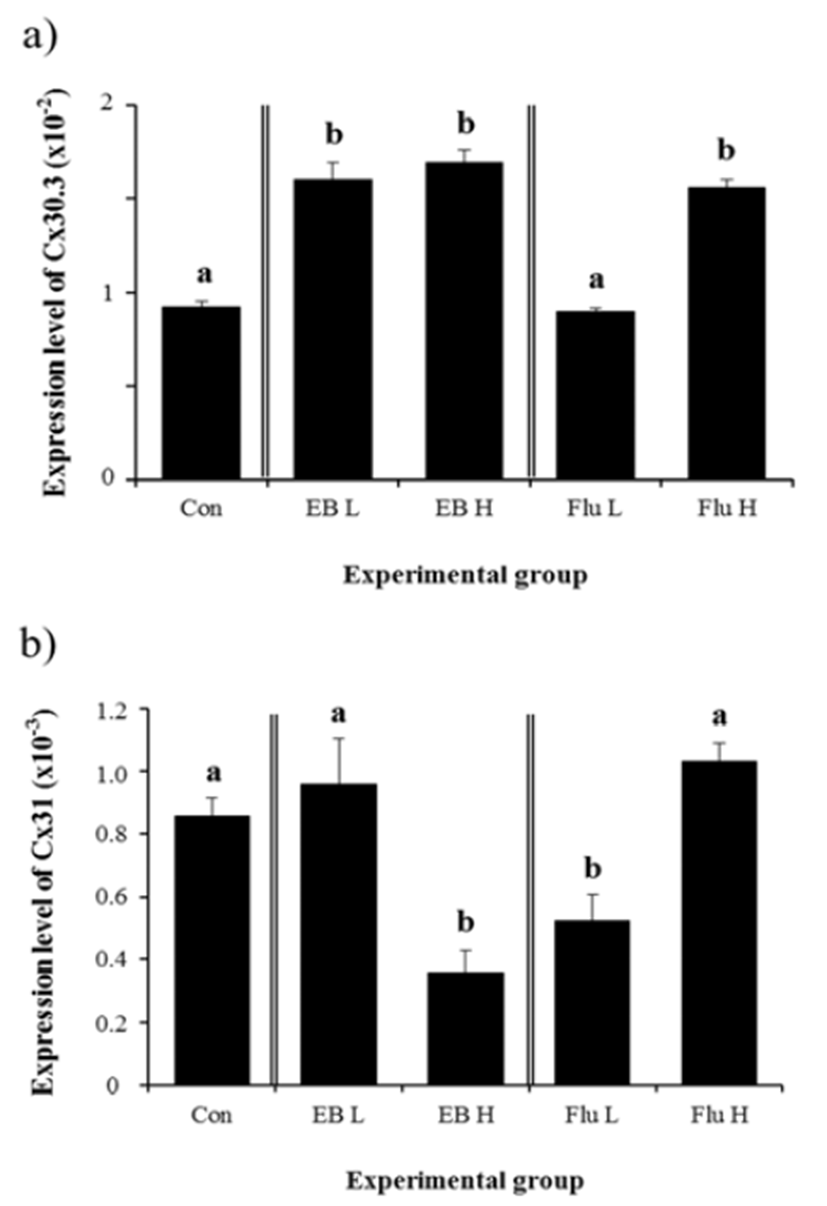
A significant increase of Cx31.1 mRNA level in the adult IS was observed with a treatment of low-dose EB at 1 week of age (Fig. 2a). However, a treatment of high-dose EB didn’t give an influence on gene expression of Cx31.1 (Fig. 2a). Expression of Cx31.1 gene in the adult IS was significantly decreased by low-dose Flu treatment at 1 week of age, while no significant change of Cx31.1 transcript level was detected with a treatment of high-dose Flu (Fig. 2a). A treatment of high-dose EB resulted in a significant decrease of Cx32 transcript abundance, but expression of Cx32 gene was not changed with low-dose EB treatment (Fig. 2b). A significant drop of Cx32 mRNA level was detected in low-dose Flu-treated group (Fig. 2b). Compared with that of control, expressional level of Cx32 gene in Flu-treated group at a high dose was significantly increased (Fig. 2b).
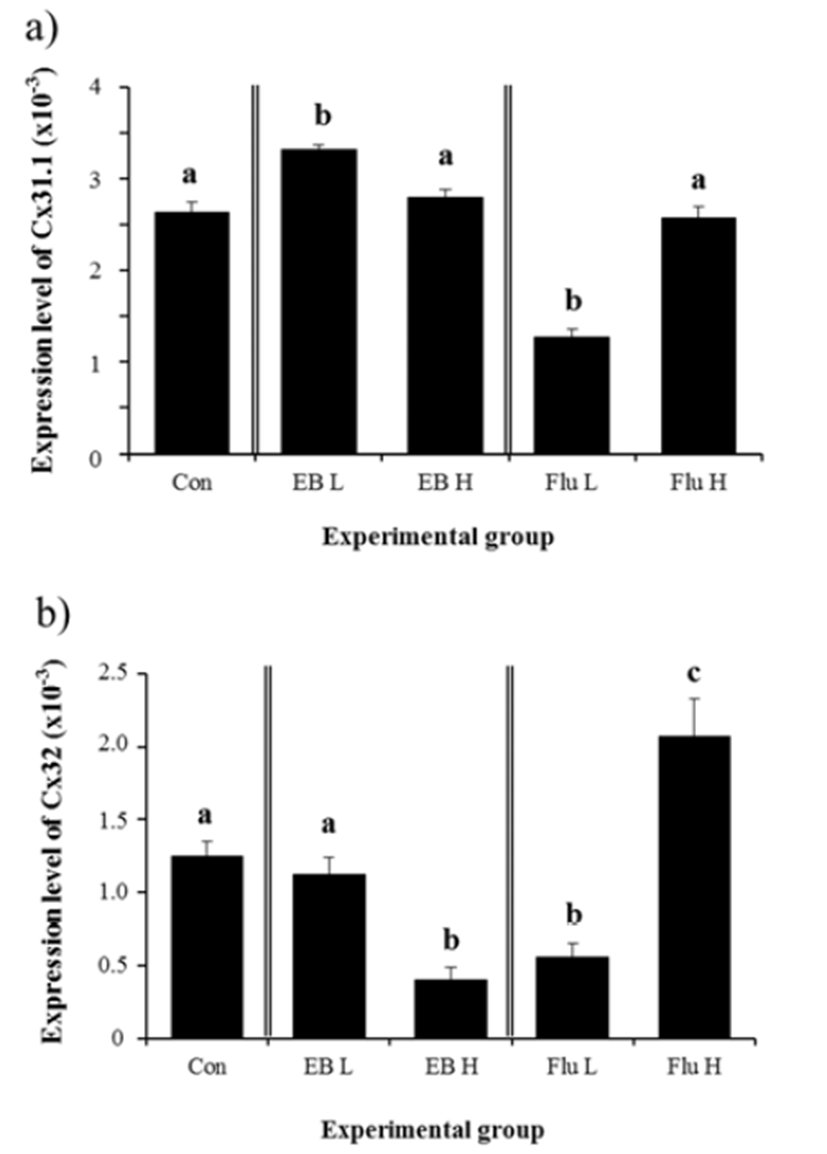
There was no significant impact on gene expression of Cx37 in the adult IS by treatments of EB at 1 week of age (Fig. 3a). A significant decrease of Cx37 mRNA level was found in low-dose Flu-treated group (Fig. 3a). A treatment of high-dose Flu at 1 week of age resulted in a significant increase of Cx37 expressional level, compared with that of control (Fig. 3a). Similarly, expression of Cx40 gene was not modulated by treatments of EB (Fig. 3b). No significant change of Cx40 gene was observed with a treatment of low-dose Flu (Fig. 3b). However, expression of Cx40 gene was significantly increased by a treatment of high-dose Flu (Fig. 3b).
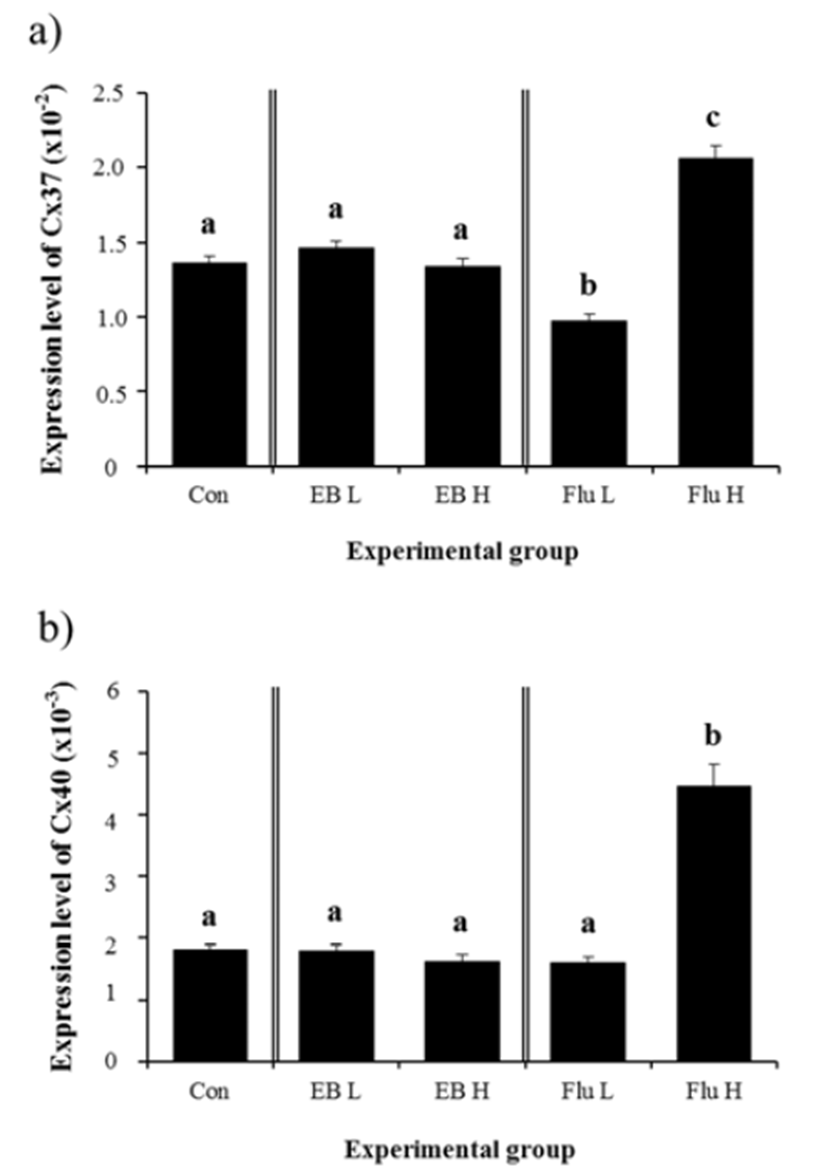
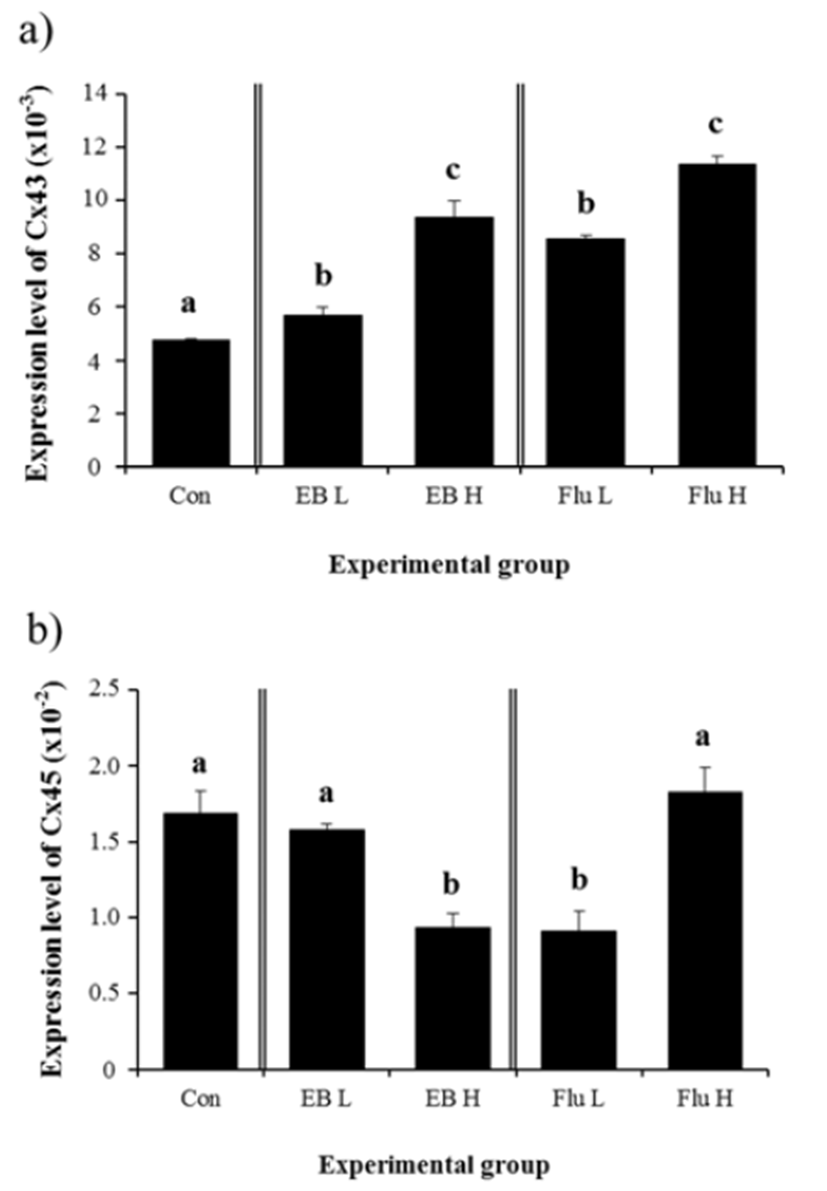
A significant increase of Cx43 mRNA level was detected in low-dose EB-treated group (Fig. 4a). Expression of Cx43 gene was further significantly increased by a treatment of high-dose EB (Fig. 4a). Similar expressional changes of Cx43 gene were observed with Flu treatment (Fig. 4a). Treatment of low-dose EB didn’t result in a significant change of Cx45 gene expression, while a significant decrease of Cx45 mRNA level was induced by a treatment of high-dose EB (Fig. 4b). In the other hand, expression of Cx45 gene was significantly decreased by a treatment of low-dose Flu, but was not affected by a treatment of highdose Flu (Fig. 4b).
DISCUSSION
Exposure to exogenous steroidal substances during the early postnatal development frequently results in disruption of systematic hormonal levels and consequently aberrant expression of genes associated with functional and structural maintenance of target tissues. The current study examined if administration of anti-androgenic or estrogenic substance at neonatal age would affect expression of Cx genes in the IS at the adult.
Neonatal exposure of bisphenol A (BPA), an environmental estrogenic compound, results in a decrease of Cx43 expression in the adult testis (Salian et al., 2009). In the present study, treatment of EB at low dose causes increases of Cx30.3, 31.1, and 43 gene expression, but not influences on the expression of other Cx genes. These findings suggest differential regulation of Cx gene expression by EB administration. Interestingly, expressional levels of Cx genes which were modulated by a treatment of EB at low dose are all increased but none is decreased. Such effects imply that a treatment of EB at low dose at neonatal age likely gives a permanent influence on Cx gene expression in the IS. At this point, it is not confirmed that expressional changes of these Cx genes are direct effects of EB action in the IS because the treatment of EB at neonatal age could give an impact on systematic balance of hormonal levels which would lead to secondary outcomes to regulate expression of Cx genes. Comparison of expressional levels of Cx gene in the IS at different recovery periods after neonatal EB exposure would provide helpful information to understand the effect of EB treatment at low dose on the expression of Cx genes.
The neonatal administration of EB at high dose results in expressional increases of Cx30.3 and 43 and expressional decreases of Cx31, 32, and 45 in the adult IS. Treatment of EB at high dose does not affect the expressional change of Cx30.3 gene, indicating that EB, regardless of the dose, stimulates transcription of Cx30.3 gene. Even though expression of Cx31, 32, and 45 genes are not influenced by treatment of EB at low dose, EB treatment at high dose leads to considerable expressional decrease of these genes, suggesting suppression of transcription of these genes by a high-dose EB. There is no expressional change of Cx37 and 40 genes by EB treatment. No influence of EB on transcriptional regulation of Cx37 and 40 genes is inferred from these observations. Intriguingly, the treatment of EB at high dose results in a further increase of Cx43 gene expression but a decrease of Cx31.1 gene expression, compared with results by EB treatment at low dose. It seems that expression of Cx31.1 and 43 genes is regulated in an opposite manner by high-dose of EB in the IS. Additional researches are suggested to examine the effect of EB on transcriptional regulation of Cx31.1 and 43 genes.
Treatment of Flu at low dose at the neonatal age leads to decreases of transcriptional levels of most Cx genes, including Cx31, 31.1, 32, 37, and 45. Interestingly, only significant increase of gene expression by a treatment of low-dose Flu is occurred in Cx43 gene, while expression of Cx30.3 and 40 genes is not affected by same treatment. Expressional pattern of Cx 31.1 by EB treatment at low dose is opposite to that by Flu treatment at low dose, implying that transcription of Cx31.1 in the IS by steroid hormones seems more susceptible than the other Cx genes. Differential regulatory effects on gene expression in hypothalamus region treated with EB or Flu at neonatal period are demonstrated from Shibutani et al. (2007) study. Expressional changes of Cx gene by an administration of Flu at high dose differ with those by a treatment of Flue at low dose. Transcript levels of Cx30.3, 32, 37, 40, and 43 are significantly increased, compared with those of control. Interestingly, expression of all Cx genes is remarkably increased, compared with those of Flu treatment at low dose, indicating a stimulatory effect of high-dose Flu treatment on expression of Cx genes. It is difficult to estimate how a treatment of Flu at high dose results in increases of Cx gene expression due to limited information available from the current study. Such effects of Flu on expression of Cx genes in the IS would be delivered from further examination with administration of various Flu doses.
Exposure to exogenous steroid hormones during neonatal period frequently results in alteration of gene expression at the adult. However, little information is available for expressional regulation of Cx genes in the IS. The present study demonstrates that EB or Flu treatment at neonatal age in male rat results in expressional modulation of most Cx genes in the adult IS. In addition, it is shown that expression of each Cx gene is differentially regulated by a type of steroid hormone and/or an amount of dose of the hormone. Moreover, the current research demonstrates a possibility of permanent and nonreversible effects induced by exposure to environmental steroid substances during the early postnatal period.